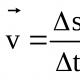Выбор читателей




Популярные статьи
Эволюция - это научная теория, которая по сути указывает на изменение видов с течением времени. Существует много разных механизмов изменения видов, но большинство из них основаны на идее естественного отбора. Эволюция посредством естественного отбора была первой научной теорией, которая предоставила доказательства изменения животных и растений со временем, а также механизм того, как это происходит.
Идея о том, что черты передаются от родителей к потомству, существует со времен древнегреческих философов. В середине 1700-х годов Кэрол Линней придумал свою систему таксономического наименования, которая группировалась по видам и подразумевала, что существует эволюционная связь между видами внутри одной группы.
В конце 1700-х годов появились первые теории, которые со временем менялись. Ученые, такие как Граф де Буффон и дедушка Чарльза Дарвина, Эразм Дарвин, предложили идею того, что виды менялись со временем, но ни один человек не мог объяснить, как и почему это происходило. Они также держали свои размышления в секрете, так как их теории были спорными относительно общепринятых религиозных взглядов той эпохи.
Жан Батист Ламарк, ученик графа де Буффона, был первым, кто публично заявил об изменении видов с течением времени. Однако, часть его теории была неверна. Ламарк предложил, что приобретенные черты передаются по наследству. Жорж Кювье смог доказать ошибочность этого утверждение. Он также имел доказательства существования видов, которые эволюционировали и вымерли.
Кювье верил в катастрофизм и считал, что эти изменения и исчезновения в природе происходили внезапно и бурно. Джеймс Хаттон и Чарльз Лайел противопоставляли аргументам Жоржа Кювье идею униформитаризма. Эта теория гласит, что изменения в природе происходят медленно и накапливаются с течением времени.
Иногда называемый «выживание наиболее приспособленных», «естественный отбор» особенно известен по книге Чарльза Дарвина «Происхождении видов» .
В книге Дарвин предложил, что виды с особенностями, наиболее подходящими для своей среды, живут достаточно долго, чтобы воспроизводить и передавать эти "удачные" черты своим потомкам. Со временем сохраняются только «наиболее приспособленные» черты вида. В конце концов, через определенный период, эти небольшие адаптации могут создавать новые виды.
В то время Чарльз Дарвин не был единственным человеком, который придумал эту идею. Альфред Рассел Уоллес также имел доказательства и пришел к похожим выводам, что и Дарвин. Они даже сотрудничали и представили совместные выводы. Вооруженные свидетельствами со всего мира благодаря многочисленным путешествиям, идеи Дарвина и Уоллеса получили положительные отзывы в научном сообществе. Партнерство закончилось, когда Дарвин опубликовал свою книгу.
Одна очень важная часть теории эволюции посредством естественного отбора - это понимание того, что виды не могут развиваться. Они могут только адаптироваться к окружающей среде. Адаптации складываются с течением времени и, в конечном итоге, приводят к эволюции вида. Это также может привести к появлению новых видов, а иногда и вымиранию более старых.
Существует множество доказательств, подтверждающих теорию эволюции. Дарвин полагался на подобные анатомии видов, чтобы связать их воедино. У него также были некоторые ископаемые доказательства, которые с течением времени демонстрировали небольшие изменения в строении тела вида, часто приводящие к рудиментарным структурам. Конечно, летопись окаменелостей неполна и имеет «недостающие звенья». С сегодняшними технологиями существует множество других доказательств эволюции. Они включает сходство эмбрионов разных видов, те же последовательности ДНК, что и у всех видов, и понимание того, как мутации ДНК работают в микроэволюции. Еще больше ископаемых доказательств было найдено со времен Дарвина, хотя в летописи окаменелостей все еще много пробелов.
Сегодня теория эволюции часто изображается в СМИ как спорный вопрос. Развитие приматов и идея о том, что люди эволюционировали от обезьян, были основной дискуссий между научными и религиозными общинами. Политики и суды решали, нужно ли школам преподавать эволюцию, или же они должны обучать альтернативным точкам зрения, таким как интеллектуальный дизайн и креационизм.
Дело Штата Теннесси против Джона Скоупса, также известное как Обезьяний процесс стало знаменитой судебной битвой за преподавание эволюции в школах. В 1925 году учитель по имени Джон Скоупс был арестован за незаконное обучение эволюции в научном классе Теннесси. Это была первое крупное судебное дело за эволюцию, и оно привлекло внимание к ранее запретной теме.
Теория эволюции часто рассматривается как главная всеобъемлющая тема, которая объединяет все темы . Сюда входят генетика, биология популяций, анатомия и физиология, а также эмбриология. В то время как эта теория сама эволюционировала и расширялась с течением времени, принципы, изложенные Дарвином в 1800-х годах, все еще сохраняются в наши дни.
Молекулярную биологию и другие.
1 / 5
✪ Синтетическая теория эволюции
✪ Эволюция - 3. Синтетическая теория эволюции - часть 1.
✪ Основные положения эволюционной теории Чарльза Дарвина. Видеоурок по биологии 9 класс
✪ Discovery - Понимание: Эволюция / Understanding: Evolution (2004)
✪ Факторы эволюции | ЕГЭ Биология | Даниил Дарвин
Вскоре после возникновения теория естественного отбора подвергалась конструктивной критике со стороны её принципиальных противников, а некоторые её элементы - и со стороны её сторонников. Большинство контраргументов против дарвинизма за первую четверть века её существования было собрано в двухтомной монографии «Дарвинизм: Критическое исследование» русским философом и публицистом Н. Я. Данилевским . Нобелевский лауреат 1908 г. И. И. Мечников , соглашаясь с Дарвином по вопросу ведущей роли естественного отбора, не разделял дарвиновскую оценку важности перенаселения для эволюции. Сам основатель теории наибольшее значение придавал контраргументу английского инженера Ф. Дженкина , который с лёгкой руки Дарвина получил название «кошмар Дженкина ».
В итоге в конце XIX - начале XX веков большинство биологов принимало концепцию эволюции, но мало кто считал, что естественный отбор является главной её движущей силой. Господствовать стали неоламаркизм , теория ортогенеза и комбинация менделевской генетики с мутационной теорией Коржинского - Де Фриза . Эту ситуацию английский биолог Джулиан Хаксли окрестил «затмением дарвинизма ru en ».
Несмотря на то что открытая Менделем дискретность наследственности устранила существенные затруднения, связанные с «кошмаром Дженкина», многие генетики отвергали дарвиновскую теорию эволюции.
Синтетическая теория в её нынешнем виде образовалась в результате переосмысления ряда положений классического дарвинизма с позиций генетики начала XX века. После переоткрытия законов Менделя (в 1901 г.), доказательства дискретной природы наследственности и особенно после создания теоретической популяционной генетики трудами Рональда Фишера , Джона Б. С. Холдейна-младшего и Сьюэла Райта , учение Дарвина приобрело прочный генетический фундамент.
Считают, что эволюционный акт состоялся, когда отбор сохранил генное сочетание, нетипичное для предшествующей истории вида. В итоге для осуществления эволюции необходимо наличие трёх процессов:
Все сторонники синтетической теории признают участие в эволюции трёх перечисленных факторов.
Важной предпосылкой для возникновения новой теории эволюции явилась книга английского генетика, математика и биохимика Дж. Б. С. Холдейна-младшего, издавшего её в 1932 году под названием «The causes of evolution ». Холдейн, создавая генетику индивидуального развития, сразу же включил новую науку в решение проблем макроэволюции.
Крупные эволюционные новшества очень часто возникают на основе неотении (сохранение ювенильных признаков у взрослого организма). Неотенией Холдейн объяснял происхождение человека («голая обезьяна»), эволюцию таких крупных таксонов , как граптолиты и фораминиферы . В 1933 году учитель Четверикова Н. К. Кольцов показал, что неотения в животном царстве широко распространена и играет важную роль в прогрессивной эволюции. Она ведет к морфологическому упрощению, но при этом сохраняется богатство генотипа .
Практически во всех историко-научных моделях 1937 год был назван годом возникновения СТЭ - в этом году появилась книга русско-американского генетика и энтомолога-систематика Ф. Г. Добржанского «Genetics and the Origin of Species ». Успех книги Добржанского определялся тем, что он был одновременно натуралистом и экспериментальным генетиком. «Двойная специализация Добржанского позволила ему первому перебросить твёрдый мост от лагеря экспериментальных биологов к лагерю натуралистов» (Э. Майр). Впервые было сформулировано важнейшее понятие об «изолирующих механизмах эволюции» - тех репродуктивных барьерах, которые отделяют генофонд одного вида от генофондов других видов. Добржанский ввёл в широкий научный оборот полузабытое уравнение Харди-Вайнберга . Он также внедрил в натуралистический материал «эффект С. Райта», полагая, что микрогеографические расы возникают под воздействием случайных изменений частот генов в малых изолятах, то есть адаптивно-нейтральным путём.
В англоязычной литературе среди создателей СТЭ чаще всего называют имена Ф. Добржанского , Дж. Хаксли, Э. Майра , Б. Ренша, Дж. Стеббинса. Это, конечно, далеко не полный список. Только из русских учёных, по меньшей мере, следовало бы назвать И. И. Шмальгаузена , Н. В. Тимофеева-Ресовского , Г. Ф. Гаузе , Н. П. Дубинина , А. Л. Тахтаджяна . Из британских ученых велика роль Дж. Б. С. Холдейна-младшего, Д. Лэка, К. Уоддингтона, Г. де-Бира. Немецкие историки среди активных создателей СТЭ называют имена Э. Баура, В. Циммермана, В. Людвига, Г. Хеберера и других.
В 1930-1940-е годы быстро произошел широкий синтез генетики и дарвинизма. Генетические идеи проникли в систематику, палеонтологию, эмбриологию, биогеографию. Термин «современный» или «эволюционный синтез» происходит из названия книги Дж. Хаксли «» (1942). Выражение «синтетическая теория эволюции» в точном приложении к данной теории впервые было использовано Дж. Симпсоном в 1949 году.
- элементарной единицей эволюции считается локальная популяция ;
- материалом для эволюции являются мутационная и рекомбинационная изменчивость;
- естественный отбор рассматривается как главная причина развития адаптаций, видообразования и происхождения надвидовых таксонов ;
- дрейф генов и принцип основателя выступают причинами формирования нейтральных признаков;
- вид есть система популяций, репродуктивно изолированных от популяций других видов, и каждый вид экологически обособлен;
- видообразование заключается в возникновении генетических изолирующих механизмов и осуществляется преимущественно в условиях географической изоляции.
Таким образом, синтетическую теорию эволюции можно охарактеризовать как теорию органической эволюции путём естественного отбора признаков, детерминированных генетически.
Активность американских создателей СТЭ была столь высока, что они быстро создали международное общество по изучению эволюции, которое в 1946 стало учредителем журнала «Evolution ». Журнал «American Naturalist » вновь вернулся к публикации работ по эволюционной тематике, делая акцент на синтезе генетики, экспериментальной и полевой биологии. В результате многочисленных и самых разнообразных исследований основные положения СТЭ прошли не только успешную проверку, но и видоизменялись, дополнялись новыми идеями.
В 1942 немецко-американский орнитолог и зоогеограф Э. Майр издал книгу «Систематика и происхождение видов», в которой была последовательно развита концепция политипического вида и генетико-географическая модель видообразования. Майр предложил принцип основателя , который в окончательной форме был им сформулирован в 1954. Если дрейф генов, как правило, дает причинное объяснение формированию нейтральных признаков во временном измерении, то принцип основателя в пространственном.
После публикации трудов Добржанского и Майра систематики получили генетическое объяснение тому, в чём они давно уже были уверены: подвиды и близкородственные виды различаются в значительной степени по адаптивно-нейтральным признакам.
Ни один из трудов по СТЭ не может сравниться с упомянутой книгой английского экспериментального биолога и натуралиста Дж. Хаксли «Evolution: The Modern synthesis » (1942 год). Труд Хаксли по объёму анализируемого материала и широте проблематики превосходит даже книгу самого Дарвина. Хаксли на протяжении многих лет держал в уме все направления в развитии эволюционной мысли, внимательно следил за развитием родственных наук и имел личный опыт генетика-экспериментатора. Видный историк биологии Провин так оценил труд Хаксли : «„Эволюция. Современный синтез“ была наиболее всесторонней по теме и документам, чем другие работы на эту тему. Книги Холдейна и Добржанского были написаны главным образом для генетиков, Майра для систематиков и Симпсона для палеонтологов. Книга Хаксли стала доминантной силой в эволюционном синтезе».
По объёму книга Хаксли не имела себе равных (645 страниц). Но самое интересное состоит в том, что все основные идеи, изложенные в книге, были очень ясно выписаны Хаксли на 20 страницах ещё в 1936, когда он послал в адрес Британской ассоциации содействия науки статью под названием «Natural selection and evolutionary progress ». В этом аспекте ни одна из публикаций по эволюционной теории, вышедшая в 1930-40-х годах, не может сравниться со статьей Хаксли. Хорошо чувствуя дух времени, Хаксли писал: «В настоящее время биология находится в фазе синтеза. До этого времени новые дисциплины работали в изоляции. Сейчас проявилась тенденция к унификации, которая является более плодотворной, чем старые односторонние взгляды на эволюцию» (1936). Ещё в трудах 1920-х годов Хаксли показал, что наследование приобретенных признаков невозможно; естественный отбор действует как фактор эволюции и как фактор стабилизации популяций и видов (эволюционный стазис); естественный отбор действует на малые и крупные мутации; географическая изоляция - важнейшее условие видообразования. Кажущаяся цель в эволюции объясняется мутациями и естественным отбором.
Основные положения статьи Хаксли 1936 года можно очень кратко изложить в такой форме:
Широкий синтез данных сравнительной анатомии, эмбриологии, биогеографии, палеонтологии с принципами генетики был осуществлен в трудах И. И. Шмальгаузена (1939), А. Л. Тахтаджяна (1943), Дж. Симпсона (1944), Б. Ренша (1947). Из этих исследований выросла теория макроэволюции . Только книга Симпсона была опубликована на английском языке и в период широкой экспансии американской биологии, чаще всего она одна упоминается среди основополагающих трудов.
Последнее высказывание, отражающее суть нейтрализма, никак не согласуется с идеологией синтетической теории эволюции, восходящей к концепции зародышевой плазмы А. Вейсмана , с которой началось развитие корпускулярной теории наследственности. Согласно взглядам Вейсмана, все факторы развития и роста находятся в половых клетках; соответственно, чтобы изменить организм, необходимо и достаточно изменить зародышевую плазму, то есть гены. В итоге теория нейтральности наследует концепцию генетического дрейфа, порожденную неодарвинизмом, но впоследствии им оставленную.
Появились новейшие теоретические разработки, позволившие ещё больше приблизить СТЭ к реально существующим фактам и явлениям, которые её первоначальная версия не могла объяснить. Достигнутые эволюционной биологией на настоящий момент рубежи отличаются от представленных ранее постулатов СТЭ:
Постулат о популяции как наименьшей эволюирующей единице остается в силе. Однако огромное количество организмов без полового процесса остается за рамками этого определения популяции, и в этом видится значительная неполнота синтетической теории эволюции.
Естественный отбор не является единственным движителем эволюции.
Эволюция далеко не всегда носит дивергентный характер.
Эволюция не обязательно идет постепенно. Не исключено, что в отдельных случаях внезапный характер могут иметь и отдельные макроэволюционные события.
Макроэволюция может идти как через микроэволюции, так и своими путями.
Сознавая недостаточность репродуктивного критерия вида, биологи все ещё не могут предложить универсального определения вида как для форм с половым процессом, так и для агамных форм.
Случайный характер мутационной изменчивости не противоречит возможности существования определенной канализированности путей эволюции, возникающей как результат прошлой истории вида. Должна стать широко известной и теория номогенеза или эволюция на основе закономерностей, выдвинутая в 1922-1923 гг. Л.С. Бергом . Его дочь Р. Л. Берг рассмотрела проблему случайности и закономерности в эволюции и пришла к заключению, что «эволюция совершается по разрешенным путям» эволюции в целом удовлетворительно объясняется этой теорией.
В качестве одного из критикуемых общих положений синтетической теории эволюции можно привести её подход к объяснению вторичного сходства, то есть близких морфологических и функциональных признаков, которые не были унаследованы, а возникли независимо в филогенетически далеких ветвях эволюции организмов.
Согласно неодарвинизму , все признаки живых существ полностью определяются генотипом и характером отбора . Поэтому параллелизм (вторичное сходство родственных существ) объясняется тем, что организмы унаследовали большое количество одинаковых генов от своего недавнего предка, а происхождение конвергентных признаков целиком приписывается действию отбора. Вместе с тем, хорошо известно, что черты сходства, развивающиеся в достаточно удалённых линиях, часто бывают неадаптивны и поэтому не могут быть правдоподобно объяснены ни естественным отбором, ни общим наследованием. Независимое возникновение одинаковых генов и их сочетаний заведомо исключается, поскольку мутации и рекомбинация - случайные процессы.
В ответ на такую критику сторонники синтетической теории могут возразить, что представления С. С. Четверикова и Р. Фишера о полной случайности мутаций в настоящее время значительно пересмотрены. Мутации случайны лишь по отношению к среде обитания, но не к существующей организации генома . Сейчас представляется вполне естественным, что разные участки ДНК обладают различной устойчивостью; соответственно, одни мутации будут возникать чаще, другие - реже. Кроме того, набор нуклеотидов весьма ограничен. Следовательно, существует вероятность независимого (и притом вполне случайного, беспричинного) появления одинаковых мутаций (вплоть до синтеза далекими друг от друга видами одного и аналогичных белков, которые не могли достаться им от общего предка). Эти и другие факторы обуславливают значительную вторичную повторяемость в структуре ДНК и могут объяснять происхождение неадаптивного сходства с позиций неодарвинизма как случайного выбора из ограниченного числа возможностей.
Другой пример - критика СТЭ сторонниками мутационной эволюции - связан с концепцией пунктуализма или «прерывистого равновесия». Пунктуализм основан на простом палеонтологическом наблюдении: продолжительность стазиса на несколько порядков превышает длительность перехода из одного фенотипического состояния в другое. Судя по имеющимся данным, это правило в общем справедливо для всей ископаемой истории многоклеточных животных и имеет достаточное количество подтверждений.
Авторы пунктуализма противопоставляют свой взгляд градуализму - представлению Дарвина о постепенной эволюции путём мелких изменений - и считают прерывистое равновесие достаточным поводом для отрицания всей синтетической теории. Столь радикальный подход вызвал дискуссию вокруг концепции прерывистого равновесия, длящуюся уже 30 лет. Большинство авторов сходится на том, что между понятиями «постепенная» и «прерывистая» имеется лишь количественная разница: длительный процесс предстает мгновенным событием, будучи изображен на сжатой временной шкале. Поэтому пунктуализм и градуализм следует рассматривать как дополнительные понятия. Кроме того, сторонники синтетической теории справедливо отмечают, что прерывистое равновесие не создает для них дополнительных трудностей: длительный стазис можно объяснять действием стабилизирующего отбора (под действием стабильных, относительно неизменных условий существования), а быстрое изменение - теорией смещающегося равновесия С. Райта для малых популяций, при резких изменениях условий существования и/или в случае прохождения вида или какой-либо его изолированной части, популяции, через бутылочное горлышко ISBN 5-03-001432-2
К концу XVIII в. в биологии накопился огромный описательный материал. Стало известно, что даже внешне очень далекие виды (например, окунь и обезьяна) в своем внутреннем строении обнаруживают черты сходства. Очень важным было установление по ископаемым остаткам того факта, что давно жившие на Земле виды животных и растений отличались от современных. Значит, не всегда виды растений и животных были такими, какие они теперь. Из практики сельского хозяйства известно, что внешний , строение и продуктивность растений и животных могут очень существенно изменяться с изменением условий их выращивания и содержания. Все эти факты заставляли наиболее прогрессивных ученых усомниться в неизменности видов.
Непререкаемый авторитет церкви был резко подорван назревавшими первыми буржуазными революциями в Европе. Философы-материалисты вступили в открытую борьбу с религией. В этой-то обстановке и возникла первая научная теория эволюции органического мира, созданная французским ученым Жаном Батистом Ламарком (1744-1829) и изложенная в его книге «Философия зоологии» (1809).
Твердая научная убежденность в изменяемости видов возникла у Ламарка на основе того нового фактического материала, который был получен биологией к концу XVIII в. Большое влияние оказали на Ламарка и французские философы-материалисты. Собственные работы по систематике растений и животных убедили его в том, что часто трудно установить резкие границы между близкими видами. В результате Ламарк приходит к своему первому выводу: виды не остаются постоянными, они медленно и постоянно изменяются.
Второй вывод состоял в утверждении факта эволюции. При рассмотрении всего животного мира в целом легко заметить, что наблюдается последовательное усложнение всей организации животных от самых низших (инфузории) до самых высших (). Это усложнение носит как бы ступенчатый характер « потому было названо Ламарком градацией. В факте градации Ламарк увидел отражение хода исторического развития органического мира. Так, Ламарк впервые в истории биологии сформулировал положение об эволюционном развитии живой природы: жизнь возникает путем самозарождения простейших живых тел из веществ неживой природы; дальнейшее историческое развитие идет по пути прогрессивного усложнения организмов, т. е. путем эволюции. Продуктом такой эволюции, хотя и с большими оговорками, Ламарк признает и человека.
Но утверждение факта эволюции - это еще не эволюционная теория. Всякая теория объясняет причины установленного факта. Поэтому и Ламарк, убедившись в историческом развитии органического мира, переходит к вопросу о причинах эволюции. Однако две взаимосвязанные стороны единого процесса эволюции (изменение отдельных видов и общий прогресс живой природы) Ламарк связывает с разными причинами. В этом - одна из слабых сторон его эволюционной теории. Как известно, в теории Дарвина и и прогресс объясняются одной общей причиной - естественным отбором.
Причину изменения видов, медленного и постепенного превращения одного вида в другой, Ламарк видел в медленном и постепенном изменении внешних условий: изменяются условия (климат, пища), а вслед за этим из поколения в поколение изменяются и виды. У организмов, лишенных нервной системы (растения, низшие животные), эти изменения возникают прямым путем. Так, у стрелолиста подводные листья лентовидные (прямое влияние водной среды), а надводные - с широкой листовой пластинкой (прямое влияние воздушной среды). У организмов с развитой нервной системой (высшие животные) влияние среды на организм осуществляется косвенным путем: изменение в условиях жизни изменяет поведение животного, его привычки; у животного по-другому начинают функционировать некоторые органы. В результате одни органы усиленно упражняются, другие, наоборот, перестают упражняться, а это влечет за собой изменение в строении этих органов и организма в целом.
Таким образом, изменения организмов, возникающие как прямым, так и косвенным путем, становятся, по Ламарку, сразу полезными, приспособительными.
Ламарк полагал, что органы развивались под влиянием усиленного упражнения: например, длинная шея и передние ноги у жирафа, широкие плавательные перепонки между пальцами у водоплавающих птиц и др. С другой стороны, органы, переставшие упражняться, постепенно становятся недоразвитыми: недоразвитые глаза у крота, недоразвитые крылья у страуса и др.
Если изменения организмов, вызванные прямым или косвенным воздействием условий среды, повторяются в целом ряду поколений, то они наследуются и становятся признаками новых видов. Так объяснял Ламарк изменение видов в природе.
В ламарковском толковании причин изменения видов есть серьезные недостатки. Так, действием упражнения или неупражнения органов нельзя объяснить изменения в таких признаках, как окраска покровов животного, густота меха и целый ряд других, к которым неприложимо понятие «упражнение» Как уже было показано в разделе « » , далеко не все изменения, возникающие у организмов под влиянием условий жизни, наследуются. А значит, не все они могут стать материалом для наследственного изменения видов.
Причину градации, т. е. прогрессивного усложнения живой природы в целом, Ламарк не смог объяснить действием условий жизни на организмы. Поэтому он совершенно произвольно допустил, что в природе существует некий изначальный, закон внутреннего стремления к совершенствованию. Согласно этим представлениям Ламарка, все живое, начиная с самозародившихся простейших, попадает под действие названного закона и в бесконечно длинном ряду поколений неизбежно прогрессивно усложняется. Нетрудно видеть, что такое объяснение по своему существу идеалистическое. Таким образом, факт прогрессивного усложнения организмов в ходе исторического развития не нашел у Ламарка материалистического объяснения.
Общая оценка роли Ж. Б. Ламарка в развитии биологии следующая:
С современной точки зрения основными доказательствами эволюции мира живых организмов являются:
единство живой природы , т.е. единые принципы клеточного строения, функционирования, наследственности и изменчивости всех живых организмов независимо от ступени их развития;
существование ископаемых переходных форм организмов, сочетающих признаки более древних и более молодых групп (свидетельствует об исторической связи разных групп организмов; пример — первоптица археоптерикс)’,
существование филогенетических (или палеонтологических) рядов, т.е. рядов ископаемых форм, связанных друг с другом в процессе эволюции и отражающих ее ход;
гомологичных органов, т.е. органов , имеющих общее строение и происхождение, но выполняющих разные функции (позволяет установить степень родства между организмами и проследить их эволюцию);
существование у организмов разных групп аналогичных органов , т.е. органов, имеющих внешнее сходство и выполняющих одинаковые функции, но имеющих разное происхождение (свидетельствует о сходных направлениях эволюции разных групп организмов под действием естественного отбора);
наличие у некоторых организмов рудиментов — органов, которые закладываются в ходе эмбрионального развития, но в дальнейшем перестают развиваться и остаются у взрослых форм в недоразвитом состоянии;
появление у отдельных организмов данного вида атавизмов — признаков, которые существовали у отдаленных предков, но были утрачены в ходе эволюции;
сходство зародышевого развития позвоночных (все многоклеточные животные развиваются из одной оплодотворенной яйцеклетки и проходят стадии дробления, бластулы, гаструлы, образования трехслойного зародыша и формирования органов из зародышевых листков, что свидетельствует о единстве их происхождения).
Биогенетический закон (Ф. Мюллер, Э. Геккель): каждая особь в индивидуальном развитии (онтогенезе) повторяет историю развития своего вида (филогенез), т.е. онтогенез есть краткое повторение филогенеза.
Синтетическая теория эволюции (современный дарвинизм) — учение об эволюции органического мира, разработанное на основе данных современной генетики, экологии и классического дарвинизма.
❖ Основные положения синтетической теории эволюции:
■ элементарный материал
для эволюции предоставляют мутации и их комбинации, создающие наследуемое гено- и фенотипическое разнообразие внутри вида;
■ основной движущий фактор
эволюции — естественный отбор как следствие борьбы за существование;
■ наименьшая
(элементарная) единица
эволюции — популяция;
■ каждая популяция эволюционирует независимо
от популяций того же вида;
■ как правило, эволюция является дивергентной
, т.е. один таксон может стать предком нескольких таксонов;
■ эволюция носит постепенный и длительный характер
и проходит как последовательная смена одной временной популяции чередой последующих временных популяций;
■ эволюция имеет ненаправленный характер
(т.е. не имеет определенной конечной цели);
■ макроэволюция на более высоком уровне, чем вид, идет путем микроэволюции; при этом макроэволюция подчиняется тем же закономерностям
, что и микроэволюция.
Уровни эволюционных преобразований:
■ микроэволюция,
■ макроэволюция.
Микроэволюция
— совокупность эволюционных процессов, протекающих в популяциях
и приводящих к изменениям их генофонда и последующему образованию новых видов.
■ Микроэволюция является основой исторического развития органического мира.
■Микроэволюционные изменения являются необходимой предпосылкой видообразования, однако они могут и не выходить за рамки данного вида.
Макроэволюция
— это совокупность процессов эволюционного преобразования на надвидовом уровне
, приводящих к возникновению систематических групп более высокого порядка, чем вид, — родов, семейств, отрядов, классов, типов и т. д.
■ Макроэволюция осуществляется по общим закономерностям, характерным для видообразования. Между макроэволюцией и микроэволюцией не существует принципиальных различий.
Отдельная особь не может быть единицей эволюции, так как ее генотип определяется в момент оплодотворения и она смертна. Вклад особи в эволюцию определяется ее наследственной изменчивостью и передачей генов потомкам. Эволюция происходит только в популяции — группе особей, которые доступны друг другу, могут скрещиваться между собой и давать жизнеспособное потомство.
Популяция
— это совокупность особей одного вида, длительно существующих на определенной территории и относительно изолированных от других особей того же вида.
■ Популяция является формой существования вида в конкретных условиях среды.
■ Популяция — наименьшая часть вида, представляющая собой элементарную единицу эволюции
.
♦Основные характеристики популяции: численность, плотность, половой и возрастной состав, генетический полиморфизм.
❖ Свойства популяций:
■ в одной популяции особи максимально сходны по признакам
(это объясняется большой вероятностью скрещивания особей внутри популяции и одинаковым давлением отбора);
■ в популяциях идет борьба за существование
и действует естественный отбор
(благодаря этому выживают и оставляют потомство лишь особи с полезными в данных условиях изменениями);
■ популяции одного вида генетически разнородны
(из-за непрерывно возникающей наследственной изменчивости);
■ популяции насыщены мутациями
и имеют широкие возможности для совершенствования существующих и выработки новых приспособлений при изменении среды;
■ популяции отличаются
друг от друга частотой проявления
тех или иных признаков
(так как в разных условиях естественному отбору подвергаются разные признаки);
■ в зонах ареала, где граничат
разные популяции одного вида, происходит обмен генами
между ними (это обеспечивает генетическое единство вида и способствует его большей изменчивости и лучшей приспособляемости к условиям обитания);
■ разные популяции одного вида находятся в относительной генетической
изоляции друг от друга;
■ вследствие этого каждая популяция эволюционирует независимо
от других популяций того же вида;
■ популяция представляет собой непрерывный поток поколений и потенциально бессмертна
.
Генофонд — совокупность генотипов всех особей популяции, вида.
❖ Закон Харди — Вайнберга (1908 г.): в больших популяциях при свободном скрещивании особей и при отсутствии мутаций, отбора и смешения с другими популяциями устанавливается равновесие, характеризуемое постоянными во времени частотами встречаемости генов, гомо- и гетерозигот , причем
p 2 + 2 pq + q 2 = l; р + q = 1,
где р — частота встречаемости доминантного гена, р 2 — частота встречаемости доминантных гомозигот, q — частота встречаемости рецессивного гена, q 2 — частота встречаемости рецессивных гомозигот, 2 pq — частота встречаемости гетерозигот.
■ Такое генотипическое равновесие возможно только в популяциях с большой численностью особей и обусловлено свободным скрещиванием между ними.
Элементарное эволюционное явление — длительное и направленное изменение генофонда популяции.
■ В условиях стойкого изменения среды в определенном направлении естественный отбор из поколения в поколение будет сохранять приспособленные фенотипы, а значит, направленно перестраивать генотипы, тем самым приводя к изменению генофонда популяции.
Элементарные факторы (или предпосылки ) эволюции — факторы, приводящие к генетической изменчивости структуры популяции (т.е. к нарушению закона Харди — Вайнберга): мутационный процесс, комбинативная изменчивость, поток генов, популяционные волны, дрейф генов, естественный отбор (случайные факторы) и различные формы изоляции (ограничивающие свободное скрещивание организмов).
Мутационный процесс в популяции обусловлен действием мутагенных факторов среды. Он идет постоянно и носит случайный и ненаправленный характер. У некоторых видов генные мутации несут от 10 до 25% особей. Большая часть мутаций снижает жизнеспособность особей или нейтральна. Однако при переходе в гетерозиготное состояние мутации могут повышать жизнестойкость потомков (наблюдается явление гетерозиса при инбридинге). Доминантные мутации сразу подпадают под действие естественного отбора. Рецессивные мутации проявляются фенотипически и подпадают под действие естественного отбора лишь через несколько поколений. Постоянное возникновение мутаций и новых комбинаций генов при скрещиваниях неизбежно вызывает наследственные изменения в популяции.
Комбинативная изменчивость усиливает влияние мутационного процесса. Как показывает опыт, жизнеспособность мутаций зависит от того, какие гены их окружают. Возникнув, отдельные мутации оказываются в соседстве с определенными генами и другими мутациями. В зависимости от своего окружения одна и та же мутация может играть как положительную, так и отрицательную роль в эволюции.
Поток (или миграция) генов — обмен генами между разными популяциями одного вида в результате свободного скрещивания их особей, происходящего при сезонных перемещениях животных в периоды размножения и в результате расселения молодняка.
Значение потока генов:
■ он увеличивает генотипическую изменчивость популяции;
■ по своему влиянию на генофонд популяции нередко превосходит эффективность мутационного процесса;
■ перемещение небольшой группы особей за пределы материнской популяции может привести к появлению новой изолированной популяции, характеризующейся значительным генотипическим однообразием (эффект основателя
).
Популяционные волны (или «волны жизни ») — это периодические изменения (колебания) численности особей в популяции, связанные с периодическими изменениями интенсивности факторов внешней среды (сменой времен года, обилием или недостатком пищи, засухами, заморозками и т.п.).
Значение популяционных волн:
■ рост численности особей влечет за собой пропорциональный рост вероятности мутаций;
■ уменьшение численности особей приводит к изменению генофонда популяции (из-за выпадения некоторых аллелей генов в результате гибели особей) — дрейфу генов.
Дрейф генов — процесс случайного ненаправленного изменения частот аллелей в популяции при небольшой ее численности.
■ Последствия дрейфа генов непредсказуемы: небольшую популяцию он может как привести к гибели, так и сделать еще более приспособленной к данной среде.
Значение дрейфа генов:
■ уменьшается доля наследственной изменчивости в популяции и возрастает ее генетическая однородность (в результате разные популяции, обитающие в сходных условиях, могут утратить свое первоначальное сходство);
■ в популяции вопреки естественному отбору может сохраняться мутантный ген, снижающий жизнеспособность особей.
Естественный отбор — это процесс преимущественного выживания и последующего размножения особей с полезными в данных условиях среды наследственными изменениями признаков, следствием которого является совершенствование адаптации и видообразование (современное определение).
❖ Основные формы естественного отбора: движущий, стабилизирующий, дизруптивный.
Движущий (или направленный ) отбор — отбор в пользу особей с полезными уклонениями от ранее установившегося в популяции среднего значения признака.
■ Особи в популяции неоднородны по фенотипу, генотипу и по норме реакции (вариационная кривая). При длительном постепенном изменении условий среды в определенном направлении преимущество получают особи с отклонениями признака от среднего значения в этом направлении. Вариационная кривая смещается или расширяется в направлении приспособления к новым условиям существования. В популяции возникают новые внутривидовые формы.
Стабилизирующий отбор — отбор в пользу особей со средним значением признака, установившегося в популяции.
■ Вследствие мутационного процесса и комбинативной изменчивости в популяции всегда появляются особи с признаками, отклоняющимися от среднего значения. При отсутствии изменений условий среды такие особи элиминируются. В результате вырабатывается относительная стойкость организации вида и его генетической структуры.
Дизруптивный (или разрывающий ) отбор — отбор, направленный против среднего, ранее установившегося в популяции, значения признака и благоприятствующий особям двух или более фенотипов, отклонившихся от промежуточной формы.
Действует при сильно изменившихся условиях внешней среды, когда основная масса особей утрачивает приспособленность к ним, а преимущества приобретают особи с крайними значениями признака. В результате этого популяция разрывается по данному признаку на несколько групп, обитающих на одной территории, что приводит к ее полиморфизму .
Полиморфизм
— существование нескольких форм по определенному признаку в одной популяции.
Дополнительные формы естественного отбора:
■ балансирующий отбор поддерживает и регулирует в популяции генетическую изменчивость без возникновения новых форм (пример: две формы двухточечной божьей коровки: красная лучше переносит зимовку и преобладает весной, черная интенсивнее размножается летом и преобладает осенью); расширяет адаптивные возможности популяции;
■ дестабилизирующий отбор: преимущества получает та популяция, в которой особи оказываются наиболее разнообразными по какому-либо признаку, значительно повышающему изменчивость популяции.
В природе определенная форма отбора редко встречается в «чистом виде». Обычно видообразование начинается с преобладания одной формы отбора, а затем ведущую роль приобретает другая форма.
Адаптация (или приспособление) — это комплекс морфологических, физиологических, поведенческих и иных особенностей особи, популяции или вида, обеспечивающий успех в конкуренции с другими особями, популяциями или видами и устойчивость к воздействию факторов окружающей среды. Адаптация — результат действия факторов эволюции.

Относительный характер адаптаций : соответствуя конкретной среде обитания, адаптации теряют свое значение при ее изменении (заяц-беляк при задержке зимы или при оттепели, ранней весной заметен на фоне пашни и деревьев; водные растения при пересыхании водоемов погибают и т.д.).
| Статьи по теме: | |
|
Функции и графики. Понятие функции. Способы задания функции Табличный способ задания функции
Что означают слова "задать функцию"? Они означают: объяснить всем... "Духота и трупный запах": детские воспоминания о Сталинградской битве Воспоминания участников сталинградской битвы читать
В начале 70-х годов поздней осенью, когда уже выпал первый снег, мне... Трубецкой дети. Биографии декабристок. Биография Екатерины
Ивановны Трубецкой
План Введение 1 Биография 2 Жена декабриста 3 Произведения,... | |