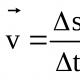Выбор читателей




Популярные статьи
Происхождение мозга Савельев Сергей Вячеславович
§ 8. Заряды мембраны нервных клеток
Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по законам электрохимическим. Нервные клетки способны получать, хранить, перерабатывать и передавать информацию при помощи специальной электрической активности. Они обладают зарядом мембраны - потенциалом покоя и могут его изменять в потенциал действия, который с высокой скоростью распространяется по телу клетки.
В основе потенциала покоя нервных клеток лежит баланс электрохимических и осмотических сил, которые действуют на клеточной границе - мембране. Мембрана клетки полупроницаема. Это означает, что через неё могут проникать далеко не все вещества. Мембрана всегда проницаема для воды, избирательно проницаема для определённых ионов и непроницаема для большинства органических соединений. Молекулы ДНК, РНК, белков и аминокислот находятся внутри клетки и не могут свободно диффундировать через мембрану. В соответствии с законами осмоса вода должна проникать в клетку. Поскольку мембрана клетки непроницаема для органических молекул, осмотическое равновесие достигнуто быть не может. Клетка должна была бы лопнуть. Этого не происходит, поскольку осмотическим силам оказывается постоянное противодействие со стороны сил совершенно другой природы.
Эти силы не осмотические, а электрохимические. Работа осмотических сил уравновешивается работой электрохимических. С одной стороны, это не позволяет клетке лопнуть, а с другой - является источником постоянного заряда мембраны нервной клетки. Внутри клетки находятся молекулы ДНК, РНК, белков, аминокислот и углеводов, которые имеют постоянный заряд. Как правило, этот заряд отрицателен и органические молекулы представляют собой набор внутриклеточных анионов (А). Их заряд уравновешивается внутри клетки положительно заряженными ионами калия (К +). Снаружи клетки основным анионом является хлор (Cl -), а катионом - натрий (Na +). В абстрактной идеальной ситуации концентрации ионов должны были бы выровняться в результате диффузии через мембрану. Однако внутриклеточные анионы неподвижны, а специальные каналы для всех подвижных ионов обычно закрыты. Более того, специализированные ионные каналы постоянно откачивают избыток натрия и хлора из клетки и закачивают внутрь внеклеточный калий. Это процесс осуществляется с затратой энергии. Она тратится на то, чтобы создать такую величину заряда мембраны, чтобы её хватило для противодействия осмотическим силам, стремящимся разрушить клетку.
В реальной клетке основные проблемы с осмотическими (гидростатическими) силами обусловлены различиями в концентрации ионов калия и натрия по обе стороны мембраны. Некоторую роль в этом
процессе играют ионы натрия и подвижность воды, свободно движущейся через мембрану клетки. Тем не менее основное значение имеет калий, поскольку его концентрационные различия максимальны. Внутри клетки калия примерно в 40 раз больше, а натрия в 9 раз меньше, чем в межклеточном пространстве, поэтому калий стремится уравновесить ситуацию, двигаясь по концентрационному градиенту из клетки, а натрий - в клетку. Поскольку концентрации этих ионов внутри и снаружи клетки известны, можно выразить эти процессы в реальных физических величинах. Работа, которую надо выполнить для предотвращения движения ионов калия из клетки по концентрационному градиенту (А 0), будет равна:
где R - газовая постоянная; Т - абсолютная температура; [К + ] - молярная концентрация калия внутри (in) и снаружи клетки (out).
Противодействовать осмотическим силам, стремящимся вывести калий из клетки, могут только электрические силы. Равновесие может быть достигнуто только при равенстве работы осмотических и электрических сил (Аэ):
Для каждого иона может быть вычислена работа, необходимая для преодоления равновесия электрических сил, возникающих вследствие разделения зарядов по обе стороны мембраны:
A 3 = FE,
где F - количество электрических зарядов в моле вещества (постоянная Фарадея); E - выраженная в вольтах разность электрических потенциалов, возникающая в результате разделения зарядов по обе стороны мембраны клетки.
Объединив формулы, легко получить известное уравнение Нернста, или равновесный (диффузионный) потенциал:

Подставив в формулу равновесия сил реальные значения при комнатной температуре, мы получим реальный заряд мембраны, который называют мембранным потенциалом. Обычно он колеблется в различных клетках от -60 до -90 мВ. При прямых измерениях зарядов мембран нервных клеток получены идентичные результаты. Обычная нервная клетка позвоночного обладает постоянным потенциалом мембраны около -75 мВ. Надо отметить, что подвижны ионы калия, натрия и хлора, поэтому при точных вычислениях мембранных потенциалов надо учитывать другие ионы.
Заряд мембраны и её способность изменять проницаемость для ионов под влиянием различных причин - уникальное эволюционное достижение. Этим свойством обладают многие клетки. Однако нервные клетки используют его для восприятия, передачи и хранения информации. Если при местном воздействии на нервную клетку возникает локальный участок с изменённым зарядом, то сигнал распространяется по мембране нервной клетки. Его обычно называют пассивным, а сам потенциал - градуальным. Это означает, что возникшее локальное возбуждение распространяется по мембране электротонически, что приводит к его постепенному затуханию. Обычно такие сигналы распространяются на небольшие расстояния, хотя у членистоногих известны нейроны, передающие такие сигналы на десятки миллиметров. Градуальные потенциалы образуют светочувствительные клетки сетчатки насекомых и позвоночных, многих периферических рецепторов и даже мотонейронов мышц стенки тела круглых червей.
Потенциал покоя есть у всех клеток на этой планете. Однако в нервной системе он является только отправной точкой для получения, хранения, обработки и передачи сигналов. Информация передаётся в нервных клетках при помощи динамического изменения заряда мембраны клетки. Это изменение может быть быстрым или медленным. Если оно происходит быстро и изменение заряда мембраны клетки превышает определённый порог, то возникает потенциал действия. Нейроны, формирующие потенциал действия, называют спайковыми. Потенциал действия отличается от градуального потенциала тем, что не затухает по мере движения от места возникновения. Причиной этого является способность мембраны активно распространять местную перезарядку мембраны, если изменение её потенциала достигнет определённого уровня. В обычной ситуации мембрана нервной клетки с потенциалом покоя -75 мВ должна изменить свой заряд до +40 мВ. Такое изменение приводит к формированию незатухающего потенциала действия, который распространяется по мембране клетки. Величину заряда мембраны, с которой начинается распространение сигнала, называют пороговой. Поскольку отростки нервных клеток достаточно хорошо изолированы глиальными клетками, а потенциал распространяется по мембране, его распространение скачкообразное (сальтаторное). В местах, где глиальные клетки плотно прилежат к нейронным мембранам, проведение происходит скачком: от одного перехвата Ранвье до другого. В перехватах мембрана отростков открыта, что позволяет осуществлять распространение сигнала путём открывания и закрывания калиевых и натриевых ионных каналов.
Принцип передачи сигнала по мембране нервной клетки довольно прост. В его основе лежит высокая плотность ионных каналов в мембране нервных клеток. Количество натриевых каналов может достигать в перехватах Ранвье 12 000 на 1 мкм 2 . Большое количество ионных каналов в мембранах нервных клеток является основой для распространения потенциалов действия. Натриевые и калиевые каналы расположены довольно плотно, что позволяет формировать локальные встречные потоки ионов при движении сигнала. Потенциал действия образуется в нейроне у аксонного холмика и начинается с открытия натриевых ионных каналов и проникновения натрия внутрь клетки. Поскольку в покое мембрана нейрона имеет заряд внутренней поверхности около -70 мВ, начинается небольшой сдвиг потенциала. Заряд мембраны локально уменьшается и постепенно доходит до нуля, а затем и до +40 мВ.
Изменение знака заряда мембраны приводит к закрыванию натриевых каналов и открыванию калиевых. Поток ионов калия восстанавливает исходный отрицательный заряд. Этот процесс называют кратковременной реверсией потенциала, или потенциалом действия. Возникнув в аксонном холмике, он распространяется по мембране клетки. Волна изменения заряда мембраны двигается с высокой скоростью, а весь процесс реверсии и восстановления занимает миллисекунды. Однако скорости движения потенциалов действия существенно различаются. На это влияют степень миелинизации, диаметр нервных волокон и многие другие факторы.
Скорости проведения потенциалов действия в нервной системе у разных видов животных различаются в десятки раз. Самые высокие скорости известны у аксонов человека и креветок (120–200 м/с), а самые низкие у актиний и медуз (0,1–0,5 м/с). По сути дела процессы передачи сигналов происходят в нервной системе со скоростью, которая отражает динамику и продолжительность жизни организма. Скорость нервных процессов определяет активность животного и внутреннее восприятие времени.
Надо отметить, что передаваемая информация кодируется частотой, последовательностью и продолжительностью активности импульсов, что обеспечивает высокую точность переносимой информации. Эти информационные сигналы различаются не только по принципам кодировки, но и по источникам - типам нервных клеток. Существует морфологическая и физиологическая классификация нейронов. По ведущим функциям нейроны физиологически подразделяются на сенсорные (афферентные), моторные, или двигательные (эфферентные), вставочные (ассоциативные, соединяющие афферентные и эфферентные) и нейросекреторные (гормональные клетки нервной системы). Эти типы не абсолютно жёсткие, а каждая клетка отчасти обладает всеми перечисленными функциями.
Из книги Микробиология: конспект лекций автора Ткаченко Ксения Викторовна2. Строение клеточной стенки и цитоплазматической мембраны Клеточная стенка – упругое ригидное образование толщиной 150–200 ангстрем. Выполняет следующие функции:1) защитную, осуществление фагоцитоза;2) регуляцию осмотического давления;3) рецепторную;4) принимает
Из книги Биология [Полный справочник для подготовки к ЕГЭ] автора Лернер Георгий Исаакович Из книги Основы психофизиологии автора Александров Юрий Из книги Проблемы лечебного голодания. Клинико-экспериментальные исследования [все четыре части!] автора Анохин Петр Кузьмич1. РЕГИСТРАЦИЯ ИМПУЛЬСНОЙ АКТИВНОСТИ НЕРВНЫХ КЛЕТОК Изучение активности нервных клеток, или нейронов, как целостных морфологических и функциональных единиц нервной системы, безусловно, остаётся базовым направлением в психофизиологии. Одним из показателей активности
Из книги Происхождение мозга автора Савельев Сергей ВячеславовичОсобенности подвижности основных нервных процессов у больных с различными психическими заболеваниями в процессе лечения их дозированным голоданием Ю. С. НИКОЛАЕВ, В. А. БРЮЗГИН, В. Б, ГУРВИЧ (Москва) В ряде предыдущих сообщений было высказано мнение, что при лечении
Из книги В поисках памяти [Возникновение новой науки о человеческой психике] автора Кандель Эрик Ричард§ 9. Синаптические контакты нервных клеток Каждый нейрон способен воспринимать и передавать информацию. Он осуществляет это в специальных участках мембраны (см. рис. I-12). От тела нейрона обычно отходят одиночные аксоны, по которым нейрон передаёт потенциалы действия или
Из книги Гены и развитие организма автора Нейфах Александр Александрович§ 10. Типы объединения нервных клеток Нервные клетки объединены в нервные системы различным образом. В простейшем случае эти элементы распределены вполне равномерно по всему телу животного или по большей его части (Anderson, 1990). Равномерное распределение нервных клеток
Из книги Мозг в электромагнитных полях автора Холодов Юрий АндреевичГлава II. Возникновение нервных клеток и мозга Причиной возникновения нервной системы стала низкая скорость получения информации о внешнем и внутреннем мире организма с донервной организацией. Его ткани состояли из клеток со сходной химической, электромагнитной и
Из книги Поведение: эволюционный подход автора Курчанов Николай Анатольевич Из книги Биологическая химия автора Лелевич Владимир Валерьянович Из книги автора2. Пол половых клеток После того как гоноциты попали в половые железы, их судьба зависит от того, какие половые клетки они должны образовать - мужские, т. е. сперматозоиды, или женские, т. е. яйца. В первом случае их путь развития называют сперматогенезом, а во втором -
Из книги автора1. Форма клеток Форма клеток зависит от их внутренней структуры и свойств клеточной оболочки и от их окружения - соседних клеток и поверхностей контакта. Так, при культивировании отдельных клеток на поверхности стекла все клетки стремятся распластаться по субстрату.
Из книги автораГлава 9. Мембраны и биохимия Электронный микроскоп показал, что биохимические реакции в живой клетке протекают с активным участием мембранных процессов. Это заключение относится и к нервной, и к глиальной клетке, и к внутриклеточным органеллам.Следует признать, что
Из книги автора7.8. Сигналы нервных клеток Нервная система возникает в ходе эволюции для анализа организмами поступающей информации. С этой целью нервные клетки используют электрические и химические сигналы. Электрические сигналы бывают двух видов.Градуальный потенциал возникает в
Из книги автораГлава 9. Биологические мембраны Клетка представляет биологическую систему, основу которой составляют мембранные структуры, отделяющие клетку от внешней среды, формирующие ее отсеки (компартменты), а также обеспечивающие поступление и удаление метаболитов, восприятие и
Из книги автораРоль медиаторов в передаче нервных импульсов Большинство синапсов в нервной системе млекопитающих является химическими. Процесс передачи сигнала в химическом синапсе осуществляется посредством освобождения нейромедиаторов из пресинаптических нервных окончаний. К

Чтобы провести сигнал от предшествующей клетки до последующей, нейрон генерирует электрические сигналы внутри себя. Твои движения глазами при чтении этого абзаца, ощущение мягкого кресла под попой, восприятие музыки из наушников и многое другое основаны на том, что внутри тебя проходят сотни миллиардов электрических сигналов. Такой сигнал может зародиться в спинном мозге и пройти до кончика пальца ноги по длинному аксону. Или может преодолеть ничтожно малое расстояние в глубинах мозга, ограничиваясь пределами интернейрона с короткими отростками. Любой нейрон, получивший сигнал, прогоняет его через свое тело и выросты, и этот сигнал имеет электрическую природу.
Еще в 1859 году ученые смогли измерить скорость, с которой передаются эти электрические сигналы. Оказалось, что электричество, передаваемое по живому аксону, принципиально отличается от электрического тока в металлах. По металлическому проводу электрический сигнал передается со скоростью, близкой к скорости света (300 000 километров в секунду), ведь в металле много свободных электронов. Однако, несмотря на эту скорость, сигнал ощутимо ослабевает, преодолевая большие расстояния. Если бы по аксонам сигналы передавались тем же способом, которым передаются в металлах, то нервный импульс, идущий от нервного окончания в коже большого пальца твоей ноги, полностью затухал бы, не достигая твоего мозга — электрическое сопротивление органической материи слишком велико, а сигнал слишком слаб.
Исследования показали, что электричество передается по аксонам намного медленнее, чем по проводам, и что в основе этой передачи лежит неизвестный ранее механизм, в результате которого сигнал распространяется со скоростью около 30 метров в секунду. Электрические сигналы, идущие по нервам, в отличие от сигналов, идущих по проводам, не ослабевают по ходу своего движения. Причина этого в том, что нервные окончания не пропускают через себя сигнал пассивно, просто позволяя имеющимся в них заряженным частицам передавать его друг другу. Они являются в каждой своей точке активным излучателем этого сигнала, ретранслируя его, и подробное описание этого механизма потребует отдельной главы. Таким образом, пожертвовав высокой скоростью проведения нервных импульсов, за счет активной передачи сигнала нейрон получает гарантию того, что возникший в большом пальце ноги сигнал достигнет спинного мозга, нисколько не ослабев.

Чтобы наблюдать прохождение электрической волны возбуждения, или потенциала действия (action potential [‘ækʃən pə’tenʃəl] ), в живой клетке, достаточно простого устройства: один конец тонкой металлической проволоки помещается на наружную поверхность аксона сенсорного нейрона кожи, а другой подводится к самописцу, чертящему линию вверх при усилении сигнала, и вниз — при ослаблении. Каждое прикосновение к коже вызывает один или несколько потенциалов действия. При возникновении каждого потенциала самописец рисует узкий длинный пик.
Потенциал действия сенсорного нейрона длится всего лишь около 0,001 секунды и включает две фазы: быстрого нарастания, достигающего пика, а затем почти столь же быстрого спада возбуждения, приводящего к исходному положению. И тут самописец сообщает неожиданный факт: все потенциалы действия, возникающие в одной и той же нервной клетке, примерно одинаковы. Это можно увидеть на картинке слева: все пики, нарисованные самописцем, имеют примерно одну и ту же форму и амплитуду независимо от того, насколько сильным или продолжительным было прикосновение к коже, их вызвавшее. Слабое поглаживание или ощутимый щипок будут переданы потенциалами действия одной и той же величины. Потенциал действия представляет собой постоянный сигнал, подчиняющийся принципу «все или ничего»: после превышения раздражителем некоего порогового значения возникает всегда примерно одинаковый сигнал, не больше и не меньше обычного. А если раздражитель меньше порогового значения, то сигнал вовсе не будет передаваться: например, можно так легко коснуться кожи кончиком пера, что это прикосновение не будет чувствоваться.
Принцип «все или ничего» в возникновении потенциала действия вызывает новые вопросы. Как сенсорный нейрон сообщает о силе раздражителя - сильное или слабое давление, яркий или тусклый свет? Как он сообщает о продолжительности действия раздражителя? Наконец, как нейроны отличают один тип сенсорной информации от другого - например, как они отличают прикосновение от боли, света, запаха или звука? И как они отличают сенсорную информацию для восприятия от моторной информации для действия?

Эволюция решила вопрос о том, как сообщить о силе раздражителя, с помощью использования одного и того же вида сигналов одной и той же величины: эта сила определяется частотой (frequency [‘friːkwənsɪ] ), с которой испускаются потенциалы действия. Слабый раздражитель, например легкое прикосновение к руке, приводит к испусканию всего двух-трех потенциалов действия в секунду, в то время как сильное давление, как при щипке или ударе по локтю, может вызвать очередь из сотни потенциалов действия в секунду. При этом продолжительность ощущения определяется продолжительностью возникновения потенциалов действия.
Используют ли нейроны разные электрические коды, сообщая мозгу, что несут информацию о разных раздражителях, таких как боль, свет или звук? Оказалось, что нет! Это удивительно, но между потенциалами действия, генерируемыми нейронами из различных сенсорных систем (например, зрительной или тактильной), разница весьма незначительна! Таким образом, характер и природа ощущения не зависят от различий в потенциалах действия (что открывает довольно захватывающую перспективу для размышлений на тему «матрицы» из одноименного фильма). Нейрон, передающий слуховую информацию, устроен точно так же, как нейрон из зрительной нервной цепи, и проводят они одни и те же потенциалы действия, одним и тем же способом. Без знания, к какой нервной цепи принадлежит конкретный нейрон, только по анализу его функционирования невозможно определить, какую информацию он несет.
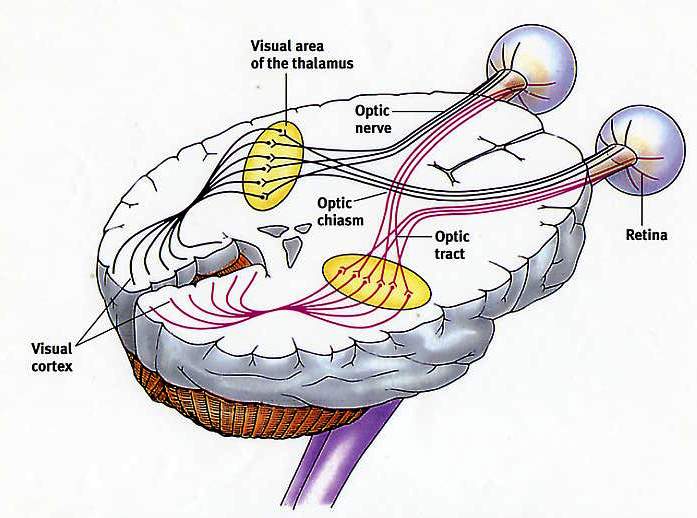
Природа передаваемой информации зависит прежде всего от типа возбуждаемых нервных волокон и специфических систем мозга, с которыми эти волокна связаны. Ощущения каждого типа передаются по своим проводящим путям, и разновидность передаваемой нейроном информации зависит именно от пути, в состав которого входит этот нейрон. В любом сенсорном проводящем пути информация передается от первого сенсорного нейрона (рецептора, реагирующего на внешний раздражитель, например прикосновение, запах или свет) к специализированным нейронам в спинном или головном мозге. Таким образом, зрительная информация отличается от слуховой лишь тем, что передается по другим проводящим путям, начинающимся в сетчатке глаза и заканчивающимся в участке мозга, который отвечает за визуальное восприятие.
Сигналы, посылаемые от моторных нейронов мозга к мышцам, также почти идентичны передаваемым по сенсорным нейронам от кожи в мозг. Они подчиняются тому же принципу «все или ничего», так же передают интенсивность сигнала с помощью частоты потенциалов действия, и так же результат сигнала зависит только от того, в какую нервную цепь включен этот нейрон. Таким образом, быстрая череда потенциалов действия, идущая по определенному проводящему пути, вызывает именно движение твоих пальцев, а не, скажем, восприятие разноцветных огней, лишь потому, что данный путь связан с мышцами рук, а не с сетчаткой глаз.

Универсальность потенциалов действия не ограничивается схожестью их проявления в разных нейронах, находящихся в пределах одного организма. Они настолько одинаковы у разных животных, что даже умудренный опытом исследователь не способен точно отличить запись потенциала действия нервного волокна кита, мыши, обезьяны или его научного руководителя. Тем не менее потенциалы действия в разных клетках не являются идентичными: небольшая разница в их амплитуде и длительности все же есть, и утверждение «все потенциалы действия одинаковы» так же неточно, как и «все бугенвиллии одинаковы».
Итак, каждый нейрон передает сигнал через свое тело и отростки одним и тем же образом. Все разнообразие информации, получаемой нами от сенсорных нейронов, все движения, которые может совершать наше тело — результат передачи единственного типа сигналов внутри нейронов. Осталась «мелочь»: понять, что же это за сигнал и как он передается.

Мы привычно отделяем все, что считаем живой природой, в том числе и себя самих, от «неживых» вещей, в том числе металлов и передающегося через них электрического тока. Тем удивительнее осознавать, что в наших телах металлы не просто присутствуют — они необходимы, без них тело не сможет существовать. Электрический ток — явление не разовое, а непрерывно возникающее в сотне миллиардов нейронов, пронизавших своими отростками все наше тело. Прямо сейчас ты можешь ощутить самые разные признаки его присутствия: то, что ты осознаешь этот текст, есть результат бесчисленных передач электрического тока. Чувство голода и удовольствие от запаха готовящейся еды, само восприятие этого запаха, прикосновение залетевшего в окно ветра к твоей коже… Перечислять можно бесконечно. И желание понять, каким же образом все это происходит, также складывается из возникающих в нейронах электрических импульсов.
Так как целью этой главы является сообщение лишь самой общей информации о прохождении нервного импульса, то здесь же необходимо рассмотреть ту среду, в которой он возникает, те условия в клетке, которые делают возможным его возникновение и передачу. Поэтому стоит начать с изучения плацдарма, на котором будут развиваться события, а именно с нейрона в состоянии покоя (dormant state [‘dɔːmənt steɪt] ).

Еще в середине прошлого века ученые нашли способ установить, в какой части нейрона существует электрический заряд. Для этого используют вольтметр (voltmeter [‘vəultˌmiːtə] ) (прибор для измерения напряжения электрического поля) с двумя электродами. Один электрод помещают внутрь нейрона, располагая его близко к клеточной мембране, а второй электрод находится в окружающей нейрон среде, с другой стороны той же мембраны. Вольтметр показывает, что с разных сторон клеточной мембраны существуют электрические заряды , отрицательный внутри клетки и положительный снаружи. Существование таких разнополюсных электрических зарядов по обе стороны мембраны создает электрическое поле, важной характеристикой которого является потенциал . Потенциал, говоря простым языком, это способность совершать работу, например работу по перетаскиванию заряженной частицы с места на место. Чем больше отрицательных зарядов накопилось по одну сторону, и чем больше положительных — по другую сторону мембраны, тем сильнее создаваемое ими электрическое поле, и тем с большей силой они способны перетаскивать туда-сюда заряженные частицы. Разницу между внешним и внутренним электрическими зарядами называют мембранным потенциалом (membrane potential [‘membreɪn pə’tenʃəl] ) покоя. Для нейрона он равен примерно 70 мВ (милливольт), то есть 70 тысячных вольта или семь сотых вольта. Для сравнения, разность потенциалов в батарейке АА равна 1,5 вольта — в 20 раз больше. То есть мембранный потенциал покоя нейрона всего лишь в 20 раз слабее, чем между клеммами батарейки АА — довольно большой, получается. Электрический потенциал существует только на мембране, и в других своих частях нейрон электрически нейтрален.
Если написать более точно, то мембранный потенциал покоя нейрона равен -70 мВ (минус семьдесят милливольт). Знак минус означает лишь то, что отрицательный заряд находится именно внутри клетки, а не снаружи, и таким образом создаваемое электрическое поле способно перетаскивать через мембрану внутрь клетки положительно заряженные ионы.
Действующие лица в создании мембранного потенциала покоя:

1 . В клеточной мембране нейрона существуют каналы, по которым несущие электрический заряд ионы могут путешествовать сквозь нее. При этом мембрана не является всего лишь пассивной «перегородкой» между внутренней средой нейрона и окружающей его межклеточной жидкостью: специальные белки, внедренные в плоть мембраны, открывают и закрывают эти каналы, и таким образом мембрана контролирует прохождение ионов — атомов, имеющих электрический заряд. Накапливая отрицательно заряженные ионы внутри клетки, нейрон увеличивает количество отрицательных зарядов внутри, тем самым приводя к увеличению положительных зарядов снаружи, и таким образом усиливается электрический потенциал. Так как протон имеет положительный заряд, а электрон отрицательный, то при избытке протонов получается положительно заряженный ион, а при избытке электронов — отрицательно заряженный. Если хочется более подробной информации об атомах и ионах, можно вернуться в . Важно понимать, что мембранный потенциал существует именно на границе клеточной мембраны, а жидкости в целом внутри и вне нейрона остаются электрически нейтральными. Ионы, для которых мембрана проницаема, остаются вблизи нее, поскольку положительные и отрицательные заряды взаимно притягиваются друг к другу. В результате снаружи мембраны образуется слой «сидящих» на ней положительных ионов, а внутри — отрицательных. Таким образом, мембрана играет роль электрической емкости, разделяющей заряды, внутри которой есть электрическое поле. Мембрана, поэтому, является природным конденсатором.
2 . отрицательно заряженные протеины , находящиеся внутри нейрона возле внутренней поверхности мембраны. Заряд протеинов всегда остается одним и тем же и является только частью общего заряда внутренней поверхности мембраны. В отличие от ионов, протеины не могут выходить из клетки и заходить в нее — для этого они слишком большие. Общий заряд меняется в зависимости от количества находящихся возле мембраны положительно заряженных ионов, концентрация которых может меняться за счет их перехода из клетки наружу, и извне вовнутрь.
3 . положительно заряженные ионы калия (К +) могут свободно перемещаться между внутренней и внешней средой, когда нейрон находится в состоянии покоя. Перемещаются они через постоянно открытые проточные калиевые каналы (flow potassium passage ), через которые могут пройти только ионы К + , и ничто другое. Проточными называются каналы, не имеющие ворот, а значит открытые при любом состоянии нейрона. Внутри клетки ионов калия гораздо больше, чем снаружи. Это происходит за счет постоянной работы натрий-калиевого насоса (про него будет рассказано ниже), поэтому в состоянии покоя нейрона ионы К + начинают перемещаться во внешнюю среду, поскольку концентрация одного и того же вещества стремится выровняться в общей системе. Если мы в бассейн с водой в одном углу выльем какое-то вещество, то его концентрация в этом углу будет очень большой, а в других частях бассейна — нулевой или очень маленькой. Однако, спустя некоторое время мы обнаружим, что концентрация этого вещества выровнялась по всему бассейну за счет броуновского движения. В этом случае говорят о «парциальном давлении» того или иного вещества, будь это жидкость или газ. Если в одном углу бассейна будет вылит спирт, то образуется большая разница в концентрации спирта между этим углом и остальным бассейном. Возникнет парциальное давление молекул спирта, и они постепенно распределятся равномерно по бассейну так, что парциальное давление исчезнет, поскольку концентрация молекул спирта везде выровняется. Таким образом, ионы К + уносят с собой положительный заряд из нейрона, уходя наружу за счет парциального давления, которое сильнее, чем сила притяжения отрицательно заряженных протеинов, в том случае, если разница в концентрации ионов внутри и снаружи клетки достаточно велика. Так как внутри остаются отрицательно заряженные протеины, то таким образом на внутренней стороне мембраны формируется отрицательный заряд. Для ясного понимания работы клеточных механизмов важно помнить, что несмотря на постоянное вытекание ионов калия из клетки, внутри нейрона их всегда больше, чем снаружи.
4 . положительно заряженные ионы натрия (Na +) находятся с внешней стороны мембраны и создают там положительный заряд. Во время фазы покоя нейрона натриевые каналы клетки закрыты , и Na + не могут пройти внутрь, а их концентрация снаружи повышается за счет работы натрий-калиевого насоса, выводящего их из нейрона.
5 . роль отрицательно заряженных ионов хлора (Cl —) и положительно заряженных ионов кальция (Ca 2+) для создания мембранного потенциала невелика, поэтому их поведение пока останется за кадром.
Формирование мембранного потенциала покоя проходит в два этапа:
Этап I . создается небольшая (-10 мВ) разница потенциалов с помощью натрий-калиевого насоса .

В отличие от других каналов мембраны, натрий-калиевый канал способен пропускать через себя и ионы натрия, и ионы калия. Причем Na + может пройти сквозь него только из клетки наружу, а К + снаружи внутрь. Один цикл работы этого канала включается в себя 4 шага:
1 . «ворота» натрий-калиевого канала открыты только с внутренней стороны мембраны, и туда заходят 3 Na +

2 . присутствие Na + внутри канала воздействует на него так, что он может частично разрушить одну молекулу АТФ (ATP ) (аденозинтрифосфата ), (adenosine triphosphate [ə’dɛnəsiːn trai’fɔsfeɪt] ) являющуюся «аккумулятором» клетки, запасающим энергию и отдающим ее при необходимости. При таком частичном разрушении, заключающемся в отщеплении от конца молекулы одной фосфатной группы PO 4 3− , выделяется энергия, которая как раз и расходуется на перенос Na + во внешнее пространство.
3 . когда канал открывается для того, чтобы Na + вышел наружу, он остается открытым, и в него попадают два иона К + — их притягивают отрицательные заряды протеинов изнутри. То, что в канале, вмещающем три иона натрия, помещается всего два иона калия, вполне логично: атом калия имеет больший диаметр.
4 . присутствие ионов калия теперь в свою очередь воздействует на канал так, что внешние «ворота» закрываются, а внутренние открываются, и К + поступают во внутреннюю среду нейрона.
Таким образом работает натрий-калиевый насос, «обменивая» три иона натрия на два иона калия. Так как электрический заряд у Na + и К + одинаковый, получается что из клетки выводится три положительных заряда, а внутрь попадает только два. За счет этого внутренний положительный заряд клеточной мембраны снижается, а внешний — увеличивается. К тому же создается разница в концентрации Na + и К + по разные стороны мембраны:
=) снаружи клетки оказывается много ионов натрия, а внутри — мало. При этом натриевые каналы закрыты, и попасть назад в клетку Na + не может, и далеко от мембраны он не уходит, так как притягивается существующим с внутренней стороны мембраны отрицательным зарядом.
=) внутри клетки много ионов калия, а вот снаружи их мало, и это приводит к вытеканию К + из клетки через открытые во время фазы покоя нейрона калиевые каналы.

Этап II формирования мембранного потенциала покоя как раз основан на этом вытекании ионов калия из нейрона. На рисунке слева показан ионный состав мембраны в начале второго этапа формирования потенциала покоя: множество К + и отрицательно заряженных протеинов (обозначенных А 4-) внутри, и облепившие мембрану снаружи Na + . Перемещаясь во внешнюю среду, ионы калия уносят из клетки свои положительные заряды, при этом суммарный заряд внутренней мембраны снижается. Так же как положительные ионы натрия, вытекшие из клетки ионы калия остаются снаружи мембраны, притягиваемые внутренним отрицательным зарядом, и внешний положительный заряд мембраны складывается из суммы зарядов Na + и К + . Несмотря на вытекание через проточные каналы, внутри клетки ионов калия всегда больше, чем снаружи.
Возникает вопрос: почему ионы калия не продолжают вытекать наружу до того момента, пока их количество внутри клетки и вне ее не станет одинаковым, то есть до тех пор, пока не исчезнет парциальное давление, создаваемое этими ионами? Причина этого заключается в том, что когда К + покидают клетку, снаружи увеличивается положительный заряд, а внутри образуется избыток отрицательного заряда. Это снижает желание ионов калия выходить из клетки, ведь наружный положительный заряд их отталкивает, а внутренний отрицательный притягивает. Поэтому через какое-то время К + перестают вытекать несмотря на то, что во внешней среде их концентрация ниже, чем во внутренней: влияние зарядов по разные стороны мембраны превышает силу парциального давления, то есть превышает стремление К + распределиться равномерно в жидкости внутри и вне нейрона. В момент достижения этого равновесия мембранный потенциал нейрона и останавливается примерно на -70 мВ.
Как только нейроном достигнут мембранный потенциал покоя, он готов для возникновения и проведения потенциала действия, про который речь пойдет в следующей цитологической главе.
Таким образом, подытожим : неравномерность распределения ионов калия и натрия по обе стороны мембраны вызвана действием двух соперничающих сил: а) силой электрического притяжения и отталкивания, и б) силой парциального давления, возникающего при разнице в концентрациях. Работа этих двух соперничающих сил протекает в условиях существования по-разному устроенных натриевых, калиевых и натриево-калиевого каналов, которые выступают в роли регуляторов действия этих сил. Калиевый канал является проточным, то есть он всегда открыт в состоянии покоя нейрона, так что ионы К + могут спокойно ходить туда-сюда под воздействием сил электрического отталкивания/притяжения и под воздействием силы, вызванные парциальным давлением, то есть разницей в концентрации этих ионов. Натриевый канал всегда закрыт в состоянии покоя нейрона, так что через них ионы Na + ходить не могут. И, наконец, натриево-калиевый канал, устроенный так, что он работает как насос, который при каждом цикле выгоняет три иона натрия наружу, и загоняет два иона калия внутрь.
Вся эта конструкция и обеспечивает возникновение мембранного потенциала покоя нейрона: т.е. состояния, при котором достигается две вещи:
а) внутри есть отрицательный заряд, а снаружи — положительный.
б) внутри много ионов К + , облепивших отрицательно заряженные части протеинов, и таким образом возникает калиевое парциальное давление — стремление ионов калия выйти наружу для выравнивания концентрации.
в) снаружи много ионов Na + , образующих отчасти пары с ионами Cl — . И таким образом возникает натриевое парциальное давление — стремление ионов натрия войти внутрь клетки для выравнивания концентрации.
В результате работы калиево-натриевого насоса мы получаем три силы, существующие на мембране: силу электрического поля и силу двух парциальных давлений. Эти силы и начинают работать, когда нейрон выходит из состояния покоя.
Ветвь наук о жизни, связанных с анатомией, физиологией, биохимией, молекулярной биологией нервной ткани и имеющих отношение к поведению и обучению называется нейронаукой. Основные составляющие нейронауки – нейробиология, психофизиология, нейрофизиология.
Нейробиология – общее название науки, занимающейся изучением на многих уровнях (начиная с молекулярного и кончая поведенческим) нервной системы и мозга как ее главного органа. Специалисты в этой области пытаются проникнуть в молекулярные, клеточные, биохимические механизмы нервных процессов. Психофизиология, или психобиология, – область знаний о биологических механизмах психических явлений. Цель ее заключается в том, чтобы понять, как из работы мозга, которая может быть представлена в виде результатов объективных измерений, возникает то, что составляет мир психических явлений. Нейрофизиология изучает тонкие механизмы работы нервных клеток мозга, называемых нейронами, природу их возбуждения и торможения.
В данной статье мы даем достаточно подробное описание механизмов взаимодействия нейронов через синапсы и детально рассматриваем участвующие в них вещества. Также рассказывается о веществах, способные повлиять на такие взаимодействия. К ним, как нетрудно догадаться, относятся психоактивные вещества, и в частности, наркотики.
Наиболее общая суть представляемой в статье картины такова. Структурной и функциональной единицей мозга является нервная клетка – нейрон. Тела нервных клеток образуют серое вещество мозга, а их отростки, из которых формируются проводящие пути и нервы, – белое вещество. Нервная ткань – совокупность нервных клеток – является характерным, хотя и не единственным компонентом нервной системы. Нейроны – специализированные клетки, способные принимать сигналы от анализаторов (органов чувств) и других нейронов, перерабатывать их в нервные импульсы и проводить эти импульсы к нервным окончаниям, контактирующим с другими нейронами или клетками органов, принимающих те или иные команды от нервной системы. Воздействие раздражителя на рецепторы трансформируется нейронами в электрические процессы. Это – общее правило для любого из органов чувств и для любых сигналов, поступающих извне в нервную систему живого существа: все воздействия из внешнего и внутреннего мира «написаны» языком электрических процессов. Перенос информации между нейронами или между нейроном и другой клеткой (рецепторная клетка органа чувств, клетка мышцы или железы и др.) осуществляется при помощи синапсов – специальных тонких щелей между конактирующими клетками. Синапсами завершаются отростки нейронов – аксоны и дендриты (они могут быть и непосредственно на теле нейрона). Импульс передается вдоль аксона или дендрита нейрона в электрической форме (мембранный потенциал). Как только импульс достигает окончания отростка нейрона (терминали), мембранный потенциал вызывает цепь событий, приводящих к выделению в синапс специфических веществ – нейротрансмиттеров (или нейромедиаторов). Они пересекают щель между контактирующими клетками и поглощаются соответствующими участками (рецепторами) на поверхности нейрона (иной клетки), расположенного по другую сторону синапса. В результате импульс передается на этот нейрон или по крайней мере на нем облегчается возникновение импульса (есть, впрочем, и такие нейротрансмиттеры, которые не облегчают, а тормозят проведение импульса на воспринявшей их клетке).
Итак, поступление сигналов в нейрон происходит через дендриты – главное место образования влияющих на нейрон возбуждающих и тормозных синапсов. Мембрана дендритов реагирует на медиатор, информация передается к телу нейрона, где комбинируется и интегрируется, и, на выходе, проходя через аксон к его окончаниям – терминалям, мембрана которых выделяет медиатор, передается следующей группе нейронов. Распространение импульсов по нейронам (возбуждение) и гашение импульсной передачи (торможение) – эти два противоположно направленных процесса лежат в основе сбалансированного, гармоничного функционирования нервной системы – предмета нейрофизиологии.
Дендриты образуют ветвящуюся систему, воспринимающую сигналы (возбуждающие и тормозные), которые суммируются в триггерной области (аксоновый холмик) мембраны нервной клетки. В нормальном (неактивном) состоянии нейрон имеет отрицательный заряд. Разность потенциалов с окружающей нейрон средой составляет –70мВ и поддерживается клеточной мембраной, обеспечивающей пониженное содержание в клетке положительных ионов натрия и повышенное содержание отрицательных ионов калия. Если в течение короткого интервала времени суммарный сигнал в триггерной области превышает –50мВ, то в нейроне развивается лавинообразный процесс, при котором за счет изменения интенсивностей потоков ионов натрия и калия между телом нейрона и окружающей средой потенциал клетки достигает +50мВ. Возникает импульс («спайк»), который по единственному аксону передается на другие нейроны. После генерации спайка наступает период, называемый рефрактерным, в течение которого клетка восстанавливает свой начальный потенциал. Этот период длится до нескольких мс.
Для передачи импульса по аксону служит миелиновая оболочка, которая примерно через каждый миллиметр прерывается узлами, называемыми перехватами Ранвье. Импульсы передаются толчками от узла к узлу, что обеспечивает, кроме минимизации энергетических затрат, значительную скорость передачи сигнала, которая может достигать 100 м/с.
Аксон на конце имеет множество ветвей, каждая из которых завершается синапсом. Все синапсы как в центральной, так и в периферической нервной системе состоят из трех элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели. Синаптическое окончание пресинаптического аксона отделяется от постсинаптического дендрита узкой синаптической щелью.
В пресинаптической мембране синтезируются медиаторы. Деполяризация пресинаптической мембраны: поступающим потенциалом действия приводит к выделению медиатора в синаптическую щель и воздействию его на постсинаптическую мембрану. При этом медиатор может оказывать как возбуждающий, так и тормозящий эффект. Нервное окончание и постсинаптическая мембрана в нервно-мышечном синапсе называются концевой, или двигательной пластинкой.
Передача сигнала в синапсе происходит химическим путем. Синапсы содержат большое количество пузырьков специального химического вещества (везикулы), которые под воздействием пришедшего сигнала лопаются и наполняют своим содержимым – медиаторами – синаптическую щель. Попавшие в синаптическую щель вещество играет роль трансмиттера, передающего возбуждение с аксона пресинаптического нейрона на дендрит постсинаптического нейрона. Таким способом постсинаптический нейрон получает сигналы по разветвленной системе своих дендритов и при достаточной совокупной мощности этих сигналов реагирует возбуждением и передачей сигнала на другие нейроны. Сила синаптической связи не является постоянной. В процессе жизнедеятельности синаптическая связь может усиливаться или ослабевать, что отражает процессы обучения и адаптации.
В зависимости от роли, которую играют нейроны в нейронной сети, различают эффекторные нейроны, передающие сигнал непосредственно исполнительным органам (например, мышцам), интернейроны (или вставочные нейроны), которые служат промежуточными звеньями по пути передачи сигнала, и рецепторные нейроны, воспринимающие сигналы от органов чувств.
Следует отметить, что нейрон не является бинарным элементом, который либо пассивен, либо возбужден. Уровень активности нейрона кодируется частотой последовательности импульсов (частотно-импульсная модуляция), которая может меняться в пределах от 1 до 100 гц.
Нейрон (нервная клетка) – это структурно-функциональная единица нервной системы. Клетка нейрона имеет сложное строение, высоко специализирована и по структуре состоит из тела (сомы) диаметром от 3 до 100 мкм, содержащего ядро и другие органеллы, и отростков. (Со строением клетки можно ознакомиться в разделе «Приложение: строение эукариотической клетки».)
Органоиды нейрона
В соме нейронов находятся ядро нейрона и ядрышко нейрона, а также хорошо развитый аппарат биосинтеза, который производит компоненты мембран, синтезирует ферменты и другие химические соединения, необходимые для специализированных функций нервных клеток. К аппарату биосинтеза в нейронах относятся тельца Ниссля – плотно прилегающие друг к другу сплюснутые цистерны гранулярного эндоплазматического ретикулума, а также хорошо выраженный комплекс Гольджи. Кроме того, сома содержит многочисленные митохондрии и элементы цитоскелета, в том числе нейрофиламенты и микротрубочки. В результате неполной деградации мембранных компонентов образуется пигмент липофусцин, накапливающийся с возрастом в ряде нейронов. В некоторых группах нейронов ствола мозга (например, в нейронах черной субстанции и голубого пятна) находится пигмент мелатонин.



Рис. Схематическое изображение двигательной нервной клетки: 1 – ядро; 2 – ядрышко; 3 – ядерная оболочка; 4 – гранулярный эндоплазматический ретикулум; 5 – комплекс Гольджи; 6 – дендриты; 7 – аксон; 8 – аксонный холмик; 9 – инициальный сегмент аксона; 10 – миелиновая оболочка; 11 – телодендрий; 12 – претерминальная и (13) терминальная части нервного волокна (аксона); 14 – пресинаптическая часть.
Ядрышко содержит большое количество РНК, покрыто тонким слоем ДНК. Существует определенная зависимость между развитием в онтогенезе ядрышка и базофильного вещества и формированием первичных поведенческих реакций у человека. Это обусловлено тем, что активность нейронов, установление контактов с другими нейронами зависят от накопления в них базофильного вещества.
Ядро нейрона окружено пористой двухслойной мембраной. Через поры происходит обмен между нуклеоплазмой и цитоплазмой. При активации нейрона ядро за счет выпячиваний увеличивает свою поверхность, что усиливает ядерно-плазматические отношения, стимулирующие функции нервной клетки. Ядро нейрона содержит генетический материал. Генетический аппарат обеспечивает дифференцировку, конечную форму клетки, а также типичные для данной клетки связи. Другой существенной функцией ядра является регуляция синтеза белка нейрона в течение всей его жизни.
Нейроны обладают рядом признаков, общих для всех клеток тела. Независимо от своего местонахождения и функций любой нейрон, как и всякая другая клетка, имеет плазматическую мембрану , определяющую границы индивидуальной клетки. Она отделяет цитоплазму клетки с включенными в нее многочисленными органоидами (ядром, аппаратом Гольджи, митохондриями и т. п.) от внеклеточной жидкости. Когда нейрон взаимодействует с другими нейронами или улавливает изменения в локальной среде, он делает это с помощью плазматической мембраны и заключенных в ней молекулярных механизмов.
Мембрана выполняет барьерную функцию, защищая клетку от поступления чужеродных веществ, и транспортную, обеспечивающую поступление в клетку необходимых для ее жизнедеятельности веществ. Через плазматическую мембрану не только регулируется поступление и выход веществ, но и осуществляется обмен информацией между клеткой и внеклеточной средой. Мембраны нервных клеток содержат множество рецепторов, активация которых приводит к повышению внутриклеточной концентрации циклического аденозинмонофосфата (цАМФ) и циклического гуанозинмонофосфата (цГМФ), регулирующих клеточный метаболизм.
Важно четко понимать, что многослойная мембрана, заключающая в себя тело (сома) нейрона, обеспечивает формирование и распространение электротонического потенциала к аксонному холмику. Нейроны способны выполнять свою информационную функцию в основном благодаря тому, что их мембрана обладает особыми свойствами. Мембрана нейрона имеет толщину 6 нм и состоит из двух слоев липидных молекул, которые своими гидрофильными концами обращены в сторону водной фазы: один слой молекул обращен внутрь, другой – кнаружи клетки. Гидрофобные концы повернуты друг к другу – внутрь мембраны. Белки мембраны встроены в двойной липидный слой и выполняют несколько функций: белки-«насосы» обеспечивают перемещение ионов и молекул против градиента концентрации в клетке; белки, встроенные в каналы, обеспечивают избирательную проницаемость мембраны; рецепторные белки распознают нужные молекулы и фиксируют их на мембране; ферменты, располагаясь на мембране, облегчают протекание химических реакций на поверхности нейрона. В ряде случаев один и тот же белок может быть и рецептором, и ферментом, и «насосом».
В отличие от большинства других клеток тела зрелые нейроны не могут делиться, и генетически обусловленные продукты любого нейрона должны обеспечивать сохранение и изменение его функций на протяжении всей его жизни .
Органеллы
Все, что находится внутри плазматической мембраны (кроме ядра), называется цитоплазмой
. Здесь содержатся цитоплазматические органеллы
, необходимые для существования нейрона и выполнения им своей работы.
Митохондрии обеспечивают клетку энергией, используя сахар и кислород для синтеза специальных высокоэнергетических молекул, расходуемых клеткой по мере надобности. Они играют важную роль в клеточном дыхании. Их больше всего у наиболее активных частей нейрона: аксонного холмика, в области синапсов. При активной деятельности нейрона количество митохондрий возрастает.
Микротрубочки – тонкие опорные структуры внутри аксона – помогают нейрону сохранять определенную форму. Сеть внутренних мембранных канальцев, с помощью которых клетка распределяет продукты, необходимые для ее функционирования, называется эндоплазматическим ретикуломом .
Существует два вида эндоплазматического ретикулума. Мембраны «шероховатого», или гранулярного, ретикулума усеяны рибосомами , необходимыми клетке для синтеза секретируемых ею белковых веществ. Обилие элементов шероховатого ретикулума в цитоплазме нейронов характеризует их как клетки с весьма интенсивной секреторной деятельностью. Белки, предназначенные только для внутриклеточного использования, синтезируются на многочисленных рибосомах, не прикрепленных к мембранам ретикулума, а находящихся в цитоплазме в свободном состоянии. Другой вид эндоплазматического ретикулума называют «гладким»
Органеллы, построенные из мембран гладкого ретикулума, упаковывают продукты, предназначенные для секреции, в «мешочки» из таких мембран для последующего переноса их к поверхности клетки, где они выводятся наружу. Гладкий эндоплазматический ретикулум называют также аппаратом Гольджи , по имени итальянца Эмилио Гольджи, который впервые разработал метод окраски этой внутренней структуры, сделавший возможным ее микроскопическое изучение. В центре цитоплазмы находится клеточное ядро . Здесь у нейронов, как и у всех клеток с ядрами, содержится генетическая информация, закодированная в химической структуре генов. В соответствии с этой информацией полностью сформированная клетка синтезирует специфические вещества, которые определяют форму, химизм и функции этой клетки.
Рибосомы располагаются, как правило, вблизи ядра и осуществляют синтез белка на матрицах тРНК. Рибосомы нейронов вступают в контакт с эндоплазматической сетью пластинчатого комплекса и образуют базофильное вещество.
Базофильное вещество (вещество Ниссля, тигроидное вещество, тигроид) – трубчатая структура, покрытая мелкими зернами, содержит РНК и участвует в синтезе белковых компонентов клетки. Длительное возбуждение нейрона приводит к исчезновению в клетке базофильного вещества, а значит, и к прекращению синтеза специфического белка. У новорожденных нейроны лобной доли коры большого мозга не имеют базофильного вещества. В то же время в структурах, обеспечивающих жизненно важные рефлексы – спинном мозге, стволе мозга, нейроны содержат большое количество базофильного вещества. Оно аксоплазматическим током из сомы клетки перемещается в аксон.
Пластинчатый комплекс (аппарат Гольджи) – органелла нейрона, окружающая ядро в виде сети. Пластинчатый комплекс участвует в синтезе нейросекреторных и других биологически активных соединений клетки. Fппарат Гольджи – система внутриклеточных мембран, каждая из которых представляет собой ряды уплощенных цистерн и секреторных пузырьков. Эту систему цитоплазматических мембран называют агранулярным ретикулумом ввиду отсутствия прикрепленных к ее цистернам и пузырькам рибосом.
Пластинчатый комплекс принимает участие в транспорте из клетки определенных веществ, в частности белков и полисахаридов. Значительная часть белков, синтезированных в рибосомах на мембранах гранулярного эндоплазматического ретикулума, поступив в пластинчатый комплекс, превращается в гликопротеины, которые упаковываются в секреторные пузырьки, а затем выделяются во внеклеточную среду. Это указывает на наличие тесной связи между пластинчатым комплексом и мембранами гранулярного эндоплазматического ретикулума.
Лизосомы и их ферменты представляют собой пузырьки, ограниченные простой мембраной и обеспечивающие фагоцитоз клетки. Они содержат набор гидролитических ферментов, способных гидролизовать вещества, попавшие в клетку. В случае гибели клетки лизосомальная мембрана разрывается и начинается процесс аутолиза – вышедшие в цитоплазму гидролазы расщепляют белки, нуклеиновые кислоты и полисахариды. Нормально функционирующая клетка надежно защищена лизосомальной мембраной от действия гидролаз, содержащихся в лизосомах.
Пигменты нейронов – меланин и липофусцин находятся в нейронах черного вещества среднего мозга, в ядрах блуждающего нерва, клетках симпатической системы.
Нейротрубочки пронизывают сому нейрона и принимают участие в хранении и передаче информации.
Нервные отростки
Нейроны сильно различаются по своей форме, по связям, которые они образуют, и способам функционирования. Наиболее очевидное отличие нейронов от других клеток заключается в разнообразии их размеров и формы. Большинство клеток тела имеет шарообразную, кубическую или пластинчатую форму. Для нейронов же характерны неправильные очертания: у них имеются отростки, часто многочисленные и разветвленные. Эти отростки - живые «провода», с помощью которых образуются нейронные цепи
.
Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ, например, нейромедиаторов. В теле нейрона выявляется развитый синтетический аппарат.
Аксон (греч. «ось») – обычно длинный отросток, приспособленный для проведения возбуждения от тела нейрона к дендритам другого нейрона. Конец аксона сильно ветвится, один аксон может контактировать с 5 тысячами нервных клеток и создавать до 10 тысяч контактов.
Специализированный участок тела клетки (чаще сомы, но иногда – дендрита), от которого отходит аксон, называется аксонным холмиком. Аксон и аксонный холмик отличаются от сомы и проксимальных участков дендритов тем, что в них нет гранулярного эндоплазматического ретикулума, свободных рибосом и комплекса Гольджи.
Дендриты – как правило, короткие и сильно разветвлённые отростки, принимающие сигнал и служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Этот термин, происходящий от греческого слова dendron – «дерево», означает, что они имеют древовидную форму.
На дендритах имеются боковые отростки (шипики), которые увеличивают их поверхность и являются местами наибольших контактов с другими нейронами.
На дендритах и на поверхности центральной части нейрона, окружающей ядро (и называемой перикарионом, или телом клетки), располагаются входные синапсы (места контактов нейронов) , образуемые аксонами других нейронов. Благодаря этому каждый нейрон оказывается звеном той или иной нейронной сети.
Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-ти тысяч) другими нейронами. Дендриты не имеют миелиновой оболочки, аксоны же имеют. Местом генерации возбуждения у большинства нейронов является аксонный холмик – образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Нервные импульсы распространяются по отросткам нейронов и передаются через синапсы (как правило, от аксонной терминали на сому или дендрит следующего нейрона). Возникновение и распространение нервного импульса, а также его синаптическая передача тесно связаны с электрическими явлениями на плазматической мембране нейрона.
На основании числа и расположения дендритов и аксона, нейроны делятся на униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентных) нейроны.
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны или ассоциативные; эта группа нейронов осуществляет связь между эфферентными и афферентными нейронами).
Афферентные нейроны (чувствительные, сенсорные нейроны) – проводят информацию об ощущении (импульс) от поверхности тела и внутренних органов в мозг.
Эфферентные нейроны (двигательные нейроны) – проводят импульс («команды») от головного и спинного мозга ко всем рабочим органам.
Вставочные нейроны (ассоциативные) – анализируют информацию и вырабатывают решения.
Вставочный нейрон передает возбуждение от афферентного (чувствительного) нейрона на эфферентные, лежит в пределах ЦНС. Тела эфферентных нейронов находятся в ЦНС или на периферии – в симпатических, парасимпатических узлах. Аксоны этих клеток продолжаются в виде нервных волокон к рабочим органам (произвольным – скелетным и непроизвольным – гладким мышцам, железам).
Нейрон выполняет трудную и тонкую работу в организме, поэтому его необходимо освободить от будничной простой работы (питание, удаление шлаков, защита от механических повреждений), что и делается клетками нейроглии (микроглия, олигодендроглия и астроглия). Клетки микроглии участвуют в образовании мозговых оболочек, клетки олигодендроглии – в образовании оболочек (милеиновых чехлов) вокруг отдельных отростков нервных клеток.
Помимо нейронов нервная ткань содержит клетки еще одного типа. Глиальные клетки окружают нервные клетки и играют вспомогательную роль. Они выполняют опорную и защитную функции и называются глиальными клетками или глией. По численности их в 10 раз больше, чем нейронов, и они занимают половину объема ЦНС. Глия не только выполняет опорные функции, но и обеспечивает многообразные метаболические процессы в нервной ткани, участвует в формировании миелиновой оболочки и способствует восстановлению нервной ткани после травм и инфекций.
Все клетки нейроглии делятся на два генетически различных вида: глиоциты (макроглия), микроглия.
К макроглии центральной нервной системы относят эпендимоциты, астроциты и олигодендроциты.
Эпендимоциты. Они образуют плотный слой клеточных элементов, выстилающих спинномозговой канал и все желудочки мозга. Выполняют пролиферативную, опорную функцию, участвуют в образовании сосудистых сплетений желудочков мозга. В сосудистых сплетениях слой эпендимы отделяет цереброспинальную жидкость от капилляров. Эпендимальные клетки желудочков мозга выполняют функцию гематоэнцефалического барьера. Некоторые эпендимоциты выполняют секреторную функцию участвуя в процессах образования цереброспинальной жидкости и выделяя различные активные вещества прямо в полость мозговых желудочков или кровь. Например, в области задней комиссуры головного мозга эпендимоциты образуют особый «субкомиссуральный орган», выделяющий секрет, возможно, участвующий в регуляции водного обмена.
Астроциты. Они образуют опорный аппарат центральной нервной системы. Различают два вида астроцитов: протоплазматические и волокнистые (фибриллярные). Между ними имеются и переходные формы. Протоплазматические астроциты лежат преимущественно в сером веществе центральной нервной системы и несут разграничительную и трофическую функции. Волокнистые астроциты располагаются главным образом в белом веществе мозга и в совокупности образуют плотную сеть – поддерживающий аппарат мозга. Отростки астроцитов на кровеносных сосудах и на поверхности мозга своими концевыми расширениями формируют периваскулярные глиальные пограничные мембраны, играющие важную роль в обмене веществ между нейронами и кровеносной системой. Эти клетки переносят питательные вещества из крови в нейроны.
Основная функция астроцитов – опорная и изоляция нейронов от внешних влияний, что необходимо для осуществления специфической деятельности нейронов.
Астроциты двух типов взаимосвязаны и образуют обширное трехмерное пространство, в которое погружены нейроны. Они часто делятся, образуя в случае повреждений центральной нервной ситемы рубцовую ткань. Олигодендроциты локализованы в сером и белом веществе. Они мельче астроцитов и содержат одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек, а само оно содержит цитоплазму с большим количеством рибосом.
Олигодендроциты (олигодендроглиоциты). Это самая многочисленная группа клеток нейроглии. Олигодендроциты окружают тела нейронов в центральной и перферической нервной системе, находятся в составе оболочек нервных волокон и в нервных окончаниях. В разных отделах нервной системы олигодендроциты имеют различную форму. Изучение методом электронной микроскопии показало, что по плотности цитоплазмы клетки олигодендроглии приближаются к нервным и отличаются от них тем, что не содержат нейрофиламентов.
Функциональное значение этих клеток очень разнообразно. Они выполняют трофическую функцию, принимая участие в обмене веществ нервных клеток. Олигодендроциты играют значительную роль в образовании оболочек вокруг отростков клеток, при этом они называются нейролеммоцитами (леммоциты – шванновские клетки). В процессе дегенерации и регенерации нервных волокон олигодендроциты выполняют еще одну очень важную функцию – они участвуют в нейронофагии (от греч. фагос – пожирающий), т.е. удаляют омертвевшие нейроны путем активного поглощения продуктов распада.
К макроглии периферической нервной системы относятся:
Клетки микроглии локализованы и в сером, и в белом веществе, но в сером веществе их больше. От каждого конца маленького продолговатого тела клетки, содержащей лизосомы и хорошо развитый аппарат Гольджи, отходит по толстому отростку. От всех его ветвей отходят более мелкие боковые веточки. При повреждении мозга эти клетки превращаются в фагоциты и, перемещаясь при помощи амебоидного движения, противостоят вторжению чужеродных частиц.
Клетки микроглии являются глиальными макрофагами и выполняют защитную функцию, принимая участие в разнообразных реакциях в ответ на повреждающие факторы. При этом клетки микроглии сначала увеличиваются в объеме, затем митотически делятся. Измененные при раздражении клетки микроглии называются зернистыми шарами.
Глиальные клетки окружают нервные клетки и в некоторых местах тесно соприкасаются с ними. Число глиальных клеток в нервной системе примерно на порядок больше числа нейронов. Особую роль глиальные клетки играют в формировании так называемых миелиновых оболочек аксонов. Миелиновые оболочки формируются у позвоночных в ЦНС за счет отростков олигодендроцитов, а на периферии – за счет так называемых шванновских клеток. Эти клетки окутывают аксоны многослойными миелиновыми «муфтами» так, что большая часть аксона оказывается покрытой ими, а открытыми остаются узкие участки между муфтами – перехваты Ранвье. Последние у таких волокон имеют особое функциональное значение.
Взаимодействие глии и нейронов. Между нейронами и глиальными клетками существуют сообщающиеся между собой щели размером 15-20 нм, так называемое интерстициальное пространство, занимающее 12-14% общего объема мозга.
Во время потенциала действия концентрация ионов калия в интерстициальном пространстве может возрастать от 3-4 ммоль/л до 10 ммоль/л, что может вызвать значительную деполяризацию нервных клеток. В результате же активного транспорта ионов калия его внеклеточная концентрация может стать ниже нормальной, что вызывает гиперполяризацию нервных клеток.
Глиальные клетки имеют высокую проницаемость для ионов калия. Когда несколько глиальных клеток деполяризуются вследствие местного повышения концентрации ионов калия, между деполяризованными и недеполяризованными клетками возникает ток, создающий вход ионам калия в деполяризованные глиальные клетки, в результате чего внеклеточная концентрация ионов калия уменьшается. Благодаря высокой проницаемости глиальных клеток для ионов калия и электрическим связям между ними глиальные клетки действуют как буфер в случае повышения внеклеточной концентрации калия.
Миелин и глиальные клетки. Многие аксоны покрыты миелиновой оболочкой, которая образована многократно закрученной мембраной глиальных клеток (шванновскими клетками). Миелин состоит преимущественно из липидов, что и придает характерный вид белому веществу головного и спинного мозга. Благодаря миелиновой оболочке скорость проведения потенциала действия по аксону увеличивается, так как ионы могут перемещаться через мембрану аксона лишь в местах, не покрытых миелином, – перехватах Ранвье . Между перехватами импульсы проводятся по миелиновой оболочке как по электрическому кабелю. Поскольку открытие канала и прохождение по нему ионов занимает какое-то время, устранение постоянного открывания каналов и ограничение их сферы действия небольшими зонами мембраны, не покрытыми миелином, ускоряет проведение импульсов по аксону примерно в 10 раз.
Только часть глиальных клеток участвует в формировании миелиновой оболочки нервов (шванновские клетки) или нервных трактов (олигодендроциты). Гораздо более многочисленные глиальные клетки (астроциты, микроглиоциты) выполняют иные функции: образуют несущий каркас нервной ткани, обеспечивают ее метаболические потребности и восстановление после травм и инфекций. Астроциты находятся вокруг нейронов, обеспечивая их механическую защиту, и, кроме того, доставляют в нейрон питательные вещества и убирают шлаки. Клетки глии обеспечивают также изоляцию одних нейронов от воздействия других. Мембрана состоит обычно из двух слоев белка, между которыми расположен слой липидов. В такую мембрану встроены разнообразные частицы. Одни из них являются частицами белка и пронизывают мембрану насквозь (интегральные белки), образуя места прохождения для ряда ионов: натрия, калия, кальция, хлора. Это, так называемые, ионные каналы. Другие частицы прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов. Это рецепторы для молекул биологически активных веществ, например медиаторов, гормонов и др.
Клеточная мембрана (плазмалемма, плазмолемма) – это тройная липопротеиновая (т.е. «жиро-белковая») оболочка, отделяющая клетку от окружающей среды и осуществлящая управляемый обмен между клеткой и окружающей её средой.
Главное в этом определении – не то, что мембрана отделяет клетку от среды, а как раз то, что она соединяет клетку с окружающей средой. Мембрана - это активная структура клетки, она постоянно работает.
Свойства мембраны:
Как видим, основное свойство мембраны – это её проницаемость по отношению к различным веществам.
Функции мембраны. Главное в работе клеточной мембраны – это обмен различными веществами между клеткой и межклеточной средой. Этому служит такое свойство мембраны как проницаемость. Кроме того, мембрана регулирует этот обмен за счёт того, что регулирует свою проницаемость.
Ещё одна важная функция мембраны – создание разности электрических потенциалов между её внутренней и наружной сторонами. За счёт этого внутри клетка имеет отрицательный электрический потенциал – потенциал покоя.
Через мембрану осуществляется также информационный обмен между клеткой и окружающей её средой. Специальные молекулярные рецепторы, расположенные на мембране, могут связываться с управляющими веществами (гормонами, медиаторами, модуляторами) и запускать в клетке биохимические реакции, приводящие к различным изменениям в работе клетки или в её структурах.
Строение мембраны. Клеточная мембрана имеет универсальное трёхслойное строение. Её срединный жировой слой является сплошным, а верхний и нижний белковые слои покрывают его в виде мозаики из отдельных белковых участков. Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами – транспортёрами и ионными каналами.

Рис. Схема трех слоёв мембраны
Живая клетка – это маленький «белково-жировой мешочек», заполненный полужидким желеобразным содержимым, которое пронизано плёнками и трубочками. Стенки этого мешочка образованы двойной жировой (липидной) плёночкой, облепленной изнутри и снаружи белками – клеточной мембраной. Поэтому говорят, что мембрана имеет трёхслойное строение: белки-жиры-белки. Внутри клетки также есть множество подобных жировых мембран, которые делят её внутреннее пространство на отсеки. Такими же мембранами окружены клеточные органеллы: ядро, митохондрии, хлоропласты. Так что мембрана – это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.
Ионные каналы мембраны. Через мембранную жировую плёнку могут проникать в клетку только жирорастворимые вещества. Это жиры, спирты, газы. Например, в эритроцитх прямо через мембрану легко проходят внутрь и наружу кислород и углекислый газ. А вот вода и водорастворимые вещества (например, ионы) просто так через мембрану не могут пройти внутрь любой клетки. Это значит, что для них нужны специальные отверстия. Но если просто сделать отверстие в жировой плёнке, то оно тут же затянется обратно. Что же делать? Выход в природе был найден: надо сделать специальные белковые транспортные структуры и протянуть их сквозь мембрану. Именно так и получаются каналы для пропускания не растворимых в жире веществ – ионные каналы мембраны клетки.
Итак, для придания своей мембране дополнительных свойства проницаемости для полярных молекул (ионов и воды) клетка синтезирует в цитоплазме специальные белки, которые затем встраиваются в мембрану. Они бывают двух типов: белки-транспортёры (например, транспортные АТФазы) и белки-каналоформеры (образователи каналов). Эти белки встраиваются в двойной жировой слой мембраны и формируют транспортные структуры в виде транспортёров или в виде ионных каналов. Через эти транспортные структуры теперь могут проходить различные водорастворимые вещества, которые по-другому проходить сквозь жировую мембранную плёнку не могут.

Рис. Мембрана нейрона и натриевые каналы
Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
Вообще, встроенные в мембрану белки ещё называются интегральными, именно потому что они как бы включаются в состав мембраны и пронизывают её насквозь. Другие белки, не интегральные, образуют как бы острова, «плавающие» по поверхности мембраны: либо по её наружной поверхности, либо по внутренней. Ведь всем известно, что жир является хорошей смазкой и скользить по нему получается легко.
Транспортные структуры мембраны – это специальные белковые структуры, встроенные в мембрану и обеспечивающие трансмембранный транспорт, т.е. перенос веществ через мембрану.
Многие вещества могут проходить сквозь клеточную мембрану самостоятельно, за счёт простой диффузии. Они «автоматически» перемещаются из зоны своей повышенной концентрации в зону пониженной концентрации, т.к. стремятся уравнять свою концентрацию по обе стороны мембраны. Так обычно ведут себя все вещества в растворах: они стремятся равномерно распределиться по всему объёму жидкости, пытаясь преодолеть разделяющую раствор на отсеки преграду. В принципе, любая молекула может пройти через липидный бислой клеточной мембраны, потому что составляющие его молекулы липидов сохраняют некоторую подвижность относительно друг друга и могут временами раздвигаться в стороны, пропуская различные вещества. Однако скорость такой пассивной диффузии, т.е. перехода вещества через мембрану из области с большей концентрацией в область с меньшей, может сильно различаться для разных веществ. Для многих веществ диффузия занимает столь длительное время, что можно говорить о практической непроницаемости для них мембраны. Скорость диффузии различных веществ через мембрану зависит главным образом от размера их молекул, электрического заряда (полярности) и их относительной растворимости в жирах.
Легче и лучше всего через мембрану пассивно проникают жирорастворимые неполярные мелкие молекулы. Так, легче всего с помощью простой диффузией проходят через мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Несколько медленнее диффундируют через липидный слой малые полярные незаряженные молекулы: СО2, NH3, Н2О, этанол, мочевина. Диффузия глицерола идёт уже значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана практически непроницаема. Таким образом, свободно проникать сквозь мембрану в клетку и обратно могут только жирорастворимые вещества, способные растворяться в жировом (липидном) слое мембраны. Транспорт других веществ через мембрану требует особых механизмов. Какие же вещества необходимо протаскивать через мембрану «насильно»? Это все полярные молекулы, не растворимые в жирах: молекулы воды, ионы (электролиты), а также более крупные молекулы питательных веществ, таких как глюкоза и аминокислоты.
Для транспорта в клетку веществ, слабо способных к диффузии через липидный слой мембраны, необходимы специальные транспортные структуры.
Виды транспортных структур мембраны:
1. Ионные каналы – специальные поры (дырочки) в мембране, образованные канальными белками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь, так и наружу.
2. Транслоказы – специальные мембранные белки, облегчающие переход вещества через мембрану за счёт своего временного связывания с диффундирующим веществом. Не требуют энергии, работают в обоих направлениях в зависимости от концентрации переносимого вещества.
3. Транспортёры – белковые структуры, насильно протаскивающие определённые вещества сквозь клеточную мембрану в определённом направлении с затратами энергии. Ионные насосы – это транспортёры ионов. По способу использования энергии для своей работы транспортёры можно разделить на «симпортные» и «антипортные». Симпортные транспортёры используют совместный транспорт в одном направлении двух веществ: одно из них должно иметь большую потенциальную энергию для движения через мембрану. Например, симпорт в клетку с помощью ионов натрия глюкозы, или симпорт ионов кальция с помощью ионов натрия. Антипортные транспортёры (обменники) используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Итак, перенос веществ через клеточную мембрану происходит различными путями.
Механизмы транспорта веществ через мембрану:
1. Простая диффузия жирорастворимых (гидрофобных) веществ через жировой слой мембраны. Это пассивный процесс под действием градиента (перепада) концентрации вещества по разные стороны мембраны. (Смотрите видео: пассивный транспорт через мембрану).
2. Неуправляемая диффузия (неуправляемый пассивный перенос) водорастворимых веществ через постоянно открытые ионные каналы мембраны.
3. Управляемая диффузия (управляемый пассивный перенос) водорастворимых веществ через управляемые ионные каналы мембраны.
4. Активный транспорт водорастворимых веществ с помощью специальных белковых транспортных структур (транспортёров) за счёт использования энергии расщепления АТФ.
Одна из самых главных транспортных структур мембраны – это фермент АТФаза. АТФазы разных видов транспортируют через мембрану ионы. Они переносят их как внутрь клетки, так и, наоборот, наружу.
Название АТФаза означает, что это фермент, нацеленный на расщепление АТФ (аденозинтрифосфатаза).
В настоящее время обраружено множество различных видов транспортных АТФаз. Они схожи между собой по строению и механизму действия, но имеют разную специализацию, т.е. каждый их вид перетаскивает через мембрану что-то своё. В настоящее время достаточно хорошо изучены Na+/K+-АТФаза, Ca2+-АТФаза, H+-АТФаза, H+,K+-АТФаза, Mg2+-АТФаза, которые обеспечивают перемещение соответственно ионов Na+, K+, Ca2+, H+, Mg2+ изолированно или сопряжённо: например, Na+ сопряжённо с К+; Н+ сопряжённо с К+.
В чём принцип работы АТФазы?
Эти ферменты расщепляют АТФ и высвобождают химическую энергию, заключённую в молекулах АТФ. Эта освобождённая энергия тратится тут же на какую-то полезную работу. Транспортные мембранные АТФазы тратят её на доставку определённого вещества на противоположную сторону мембраны «силой». Различные АТФазы, встроенные в мембрану, выполняет функцию переносчиков для различных веществ и являются, таким образом, молекулярными транспортёрами, «насильно» переносящими вещества сквозь мембрану. Такой перенос называется активным транспортом.
Самой главной мембранной АТФазой по праву можно считать Na,K-АТФазу (натрий-калиевую аденозинтрифосфатазу).
Na,K-АТФаза образует в мембране «ионный натрий-калиевый насос» , который разносит по разные стороны мембраны ионы Na+ и K+. Важно понять, что этот насос работает как обменник. На внутренней стороне мембраны активный центр фермента (АТФазы) захватывает 3 иона натрия и выбрасывает их уже на внешней стороне. А выбросив ионы натрия наружу, АТФаза на их место захватывает снаружи 2 иона калия. Затем фермент выворачивается внутрь клетки и перемещает ионы калия на внутреннюю сторону мембраны. Там он отпускает их, а вместо них опять захватывает 3 иона натрия.
При этом следует помнить, что, как истинный фермент, Na,K-АТФаза параллельно расщепляет АТФ, получая от этого энергию на свою транспортную деятельность. Далее цикл повторяется.
Мембранный потенциал покоя. Зачем нам нужно знать, что такое потенциал покоя? Что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку»?
На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка – нейрон.
А в основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы – это понять, каким образом перемещаются электрические заряды на нервных клетках к как за счёт этого на мембране появляется электрический потенцила. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов – формирование потенциала покоя.
В норме, когда клетка готова к работе, у неё уже есть электрический заряд на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя – это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет –70 мВ (милливольт).
«Потенциал» – это возможность, он сродни понятию «потенция». Электрический потенциал мембраны – это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы – ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (–), а остальные – положительно (+).
Имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами – химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках – это поток ионов, а не электронов, как в проводах. Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи.
То есть снаружи вокруг клетки будут преобладать положительно заряженные ионы, а внутри – отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна.
Сущность потенциала покоя – это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны, (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны. В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях – и тогда потенциал покоя нарушается и смещается в сторону нуля. Это называется деполяризацией мембраны, т.е. уменьшением полярности.
2. Поведение ионов калия и натрия. Ионы калия и натрия по-разному перемещаются через мембрану:
1) Через ионные насосы-обменники калий затаскивается в клетку, а натрий выводится из клетки.
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий «хочет» войти в клетку, но «не может», т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы.
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне - 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя. Наши клетки только снаружи положительные, а внутри они весьма отрицательные, и в них существует избыток отрицательных частиц – анионов и недостаток положительных частиц – катионов. Внутренняя электроотрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а наоборот – из-за потери некоторого количества положительных частиц (катионов). И поэтому сущность нашего рассказа будет заключаться не в том, что мы объясним, откуда берутся отрицательные частицы в клетке, а в том, что мы объясним, каким образом в нейронах получается дефицит положительно заряженных ионов – катионов.
Куда же деваются из клетки положительно заряженные частицы? Напомним, что это ионы натрия – Na+ и калия - K+.
А всё дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют «собственный» натрий клетки на наружный «чужой» калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Чтобы легче было запомнить, образно можно сказать так: «Клетка любит калий!» (Хотя об истинной любви здесь не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его и так полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Кстати, интересно, что клетка не рождается с потенциалом покоя в готовом виде. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от –10 до –70мВ, т.е. их мембрана становится более электроотрицательной, она поляризуется в процессе дифференцировки.
Образно говоря, можно сказать, что создавая потенциал покоя, клетка «заряжается любовью» к калию и любовью калия к свободе. Как ни странно, но результат этих двух видов любви - пустота!
Именно пустота, создаёт в клетке отрицательный электрический заряд – потенциал покоя. Точнее, отрицательный потенциал создают пустые места, оставшиеся от убежавшего из клетки калия.
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Электрический потенциал – возможность затягивать внутрь клетки положительно заряженные частицы (ионы).
2. Ионный натриевый потенциал – возможность затягивать внутрь клетки ионы натрия (и именно натрия, а не какие-нибудь другие).
3. Ионный калиевый потенциал – возможновть выталкивать из клетки ионы калия (и именно калия, а не какие-нибудь другие).
Итак, результат деятельности мембранных ионных насосов-обменников таков:
1. Дефицит натрия (Na+) в клетке.
2. Избыток калия (K+) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь «полезет» этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выравнять свою концентрацию во всём объёме раствора. И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
Тут ещё важно понять, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов в клетке надо по-отдельности сравнивать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно - концентрацию калия внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это часто делается в учебниках.
По закону выравнивания концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците. А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
А теперь – самое главное. Мы должны перейти от движения химических частиц к движению электрических зарядов.
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и положительные заряды. На их месте в клетке остаются отрицательные заряды. Это и есть мембранный потенциал покоя.
Мембранный потенциал покоя – это дефицит положительных зарядов внутри клетки, образовавшийся за счёт утечки из клетки положительных ионов калия.
Потенциал покоя – отрицательный со стороны клетки и состоит как бы из двух частей.
1. Первая часть – это примерно –10мВ, которые получаются от неравносторонней работы мембранного насоса-обменника (ведь он больше выкачивает Na+, чем закачивает обратно с K+).
2. Вторая часть – это утекающий всё время из клетки калий, утаскивающий положительные заряды из клетки. Он дает большую часть мембранного потенциала, доводя его до –70 милливольт.
Калий перестанет выходить из клетки (точнее, его вход и выход сравняются) только при уровне электроотрицательности клетки в –90мВ. Но этому мешает постоянно подтекающий в клетку натрий, который тащит с собой свои положительные заряды. И в клетке поддерживается равновесное состояние на уровне –70мВ.
Так что всё дело в натрий-калиевом мембранном насосе-обменнике и последующем вытекании из клетки «лишнего» калия. За счёт потери положительных зарядов при этом вытекании внутри клетки нарастает электроотрицательность. Она-то и есть «мембранный потенциал покоя». Он измеряется внутри клетки и составляет обычно –70 мВ.
Локальный потенциал. Локальный потенциал (ЛП) – это местное нераспространяющееся подпороговое возбуждение, существующее в пределах от потенциала покоя (–70 мВ) до критического уровня деполяризации (–50 мВ).
В случае превышения критического уровня деполяризации локальный потенциал переходит в потенциал действия и порождает нервный импульс.
Критический уровень деполяризации (КУД) – это такой уровень электрического потенциала мембраны возбудимой клетки, от которого локальный потенциал переходит в потенциал действия. В основе этого явления лежит самонарастающее открытие потенциал-управляемых ионных каналов для натрия под действием нарастающей деполяризации.
КУД обычно составляет -50 мВ, но бывает разным у разных нейронов и может меняться при изменении возбудимости нейрона. Чем ближе КУД к потенциалу покоя (–70 мВ) и, наоборот, чем ближе потенциал покоя к КУД, тем более возбудимым является нейрон.
Важно понять то, что процесс рождения локального потенциала начинается с открытия ионных каналов. Открытие ионных каналов – это самое главное. Их нужно открыть для того, чтобы в клетку пошёл поток ионов и принёс в неё электрические заряды. Эти ионные электрические заряды как раз и вызывают смещение электрического потенциала мембраны вверх или вниз, т.е. локальный потенциал.
Если открываются ионные каналы для натрия (Na+), то в клетку вместе с ионами натрия попадают положительные заряды, и её потенциал смещается вверх в сторону нуля. Это – деполяризация (уменьшение электроотрицательности нервной клетки), и так рождается возбуждающий локальный потенциал. Можно сказать, что возбуждающие локальные потенциалы порождаются натриевыми ионными каналами, когда они открываются.
Образно можно сказать и так: «каналы открываются – потенциал рождается».
Если открываются ионные каналы для хлора (Cl–), то в клетку вместе с ионами хлора попадают отрицательные заряды, и её потенциал смещается вниз ниже потенциала покоя. Это гиперполяризация, и таким способом рождается тормозный локальный потенциал. Можно сказать, что тормозные локальные потенциалы порождаются хлорными ионными каналами.
Существует также ещё один механизм формирования тормозных локальных потенциалов – за счёт открытия дополнительных ионных каналов для калия (К+). В этом случае из клетки через них начинают выходить «лишние» порции ионов калия, они выносят положительные заряды и увеличивают электроотрицательность клетки, т.е. вызывают её гиперполяризацию. Таким образом, можно сказать, что тормозные локальные потенциалы порождаются дополнительными калиевыми ионными каналами.
Как видно, всё очень просто, главное – открыть нужные ионные каналы. Стимул-управляемые ионные каналы открываются раздражителем (стимулом). Хемо-управляемые ионные каналы открываются медиатором (возбуждающим или тормозным). Точнее, в зависимости от того, на какие каналы (натриевые, калиевые или хлорные) будет действовать медиатор, таков будет и локальный потенциал – возбуждающий или тормозный. А медиатор как для возбуждающих локальных потенциалов, так и для тормозных, может быть одним и тем же, тут важно, какие ионные каналы будут связываться с ним своими молекулярными рецепторами – натриевые, калиевые или хлорные.
Виды локальных потенциалов:
1. Рецепторный. Возникает на рецепторных клетках или рецепторных окончаниях нейронов под действием стимула (раздражителя).
2. Генераторный. Возникает на сенсорных афферентных нейронах (на их нервных окончаниях, перехватах Ранвье или аксонных холмиках) под действием медиаторов, которые выделили сенсорные клеточные рецепторы. Генераторный потенциал превращается в потенциал действия и нервный импульс при достижении им критического уровня деполяризации, т.е. он генерирует (порождает) нервный импульс.
3. Возбуждающий постсинаптический потенциал (ВПСП). Возникает на постсинаптической мембране синапса, т.е. он отражает передачу возбуждения от одного нейрона к другому. Он вызывает деполяризацию мембраны. Но обычно требуется целая серия ВПСП для того, чтобы родился нервный импульс, т.к. величины единичного ВПСП совершенно недостаточно для того, чтобы достичь критического уровня деполяризации.
4. Тормозный постсинаптический потенциал (ТПСП). Возникает на постсинаптической мембране синапса, но только не возбуждает её, а, наоборот, тормозит. Соотвтетственно, эта постсинаптическая мембрана входит в состав тормозного синапса, а не возбуждающего. ТПСП вызывает гиперполяризацию мембраны, т.е. сдвигает потенциал покоя вниз от нуля. Используются два механизма создания ТПСП: 1) «хлорный» – открытие ионных каналов для хлора (Cl–), через которые в клетку входят ионы хлора и увеличивают её отрицательность, 2) «калиевый» – открытие ионных каналов для калия (К+), через которые выходят ионы калия, что также увеличивает отрицательность в клетке.
5. Пейсмекерные потенциалы – это эндогенные близкие к синусоидальным периодические колебания мембранного потенциала с частотой 0,1-10 Гц и амплитудой 5-10 мВ. Они генерируются нейронами-пейсмекерами (водителями ритма) самостоятельно, без внешнего воздействия. Пейсмекерные локальные потенциалы обеспечивают периодическое достижение нейроном-пейсмекером критического уровня деполяризации и спонтанную (самопроизвольную) генерацию им потенциалов действия и, соответственно, нервных импульсов.
Локальные потенциалы возникюет на сенсорных рецепторах, на рецепторных окончаниях нейронов и на постсинаптических мембранах синапсов.
Потенциал действия. Строго говоря, движущееся по нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия.
Нервный импульс – это движущаяся волна изменений в состоянии мембраны, включающая в себя структурные изменения (открытие и закрытие ионных каналов), химические (изменяющиеся потоки ионов) и электрические (деполяризацию, позитивную поляризацию и реполяризацию).
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин «потенциал действия». Хотя потенциал действия – это только электрический компонент нервного импульса.
Потенциал действия – это электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от –70 до +30 мВ и обратно).
Потенциалом действия называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения. По сути своей он действия представляет электрический разряд – быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.

Рис. Нервная клетка и потенциал действия
Схема распределения зарядов по разные стороны мембраны возбудимой клетки в спокойном состоянии (A) и при возникновении потенциала действия - деполяризация (B).
Нервные импульсы рождаются на нейронах всего в нескольких местах:
1. Аксонный холмик – главный породитель нервных импульсов. Аксонный холмик – это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интергрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) – и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим «начинаниям». Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они «перебиваются» потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона. Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита). Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса. В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения можно рассматривать как единый электрохимический процесс распространяющегося динамичного возбуждения с трех сторон, на трех уровнях:
Три стороны процесса распространяющегося возбуждения:
1. Потенциал действия (ПД). Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию. Сначала деполяризация идет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением. Говоря образно, деполяризация так разгоняется, что перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация, т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия.

Восходящая ветвь графика:
Нисходящая ветвь графика:
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки. Образно можно сказать, что заряженные ионы – это и есть создатели электрических потенциалов в нервных клетках. Для многих людей звучит странно утверждение, что вода не проводит электрический ток. Но на самом деле это так. Сама по себе вода является диэлектриком, а не проводником. В воде электрический ток обеспечивают не электроны, как в металлических проводах, а заряженные ионы: положительные катионы и отрицательные анионы. В живых клетках основную «электрическую работу» выполняют катионы, так как они более подвижны. Электрические токи в клетках – это потоки ионов.
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками. Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы. Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы.
На молекулярно-структурном уровне происходит открытие мембранных натриевых каналов. Сначала этот процесс идет пропорционально силе внешнего воздействия, а затем становится просто «неудержимым» и массовым. Открытие каналов обеспечивает вход натрия в клетку и вызывает деполяризацию. Затем, примерно через 2-5 миллисекунд, происходит их автоматическое закрытие. Это закрытие каналов резко обрывает движение ионов натрия внутрь клетки, и, следовательно, обрывает нарастание электрического потенциала. Рост потенциала прекращается, и на графике мы видим спайк. Это вершина кривой на графике, дальше процесс пойдет уже в обратном направлении. Конечно, очень интересно разобраться в том, что натриевые каналы имеют двое ворот, и открываются они активационными воротами, а закрываются инактивационными, но это следует обсуждать ранее, в теме «Возбуждение». Мы на этом останавливаться не будем.
Параллельно в открытием натриевых каналов с небольшим отставанием во времени идет нарастающее открытие калиевых каналов. Они медлительные по сравнению с натриевыми. Открытие дополнительных калиевых каналов усиливает выход положительных ионов калия из клетки. Выход калия противодействует «натриевой» деполяризации и вызывает восстановление полярности (восстановление электроотрицательности). Но натриевые каналы опережают калиевые, они срабатывают примерно в 10 раз быстрее. Поэтому входящий поток положительных ионов натрия в клетку опережает компенсирующий выход ионов калия. И поэтому деполяризация развивается опережающими темпами по сравнению с противодействующей ей поляризацией, вызванной утечкой ионов калия. Вот почему, пока натриевые каналы не закроются, восстановление поляризации не начнется.
Для того чтобы перейти к пониманию смысла динамичного процесса возбуждения, т.е. к пониманию его распространения вдоль мембраны, надо представить себе, что описанные нами выше процессы захватывают сначала ближайшие, а затем все новые, все более и более отдаленные участки мембраны, пока не пробегут по всей мембране полностью. Если вы видели «живую волну», которую устраивают болельщики на стадионе за счет вставания и приседания, то вам легко будет представить себе мембранную волну возбуждения, которая образуется за счет последовательного протекания в соседних участках трансмембранных ионных токов.
В качестве аналогии или метафоры, которая может наглядно передать смысл распространяющегося возбуждения, можно взять обрае пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
Можно ли «погасить» волну распространяющегося возбуждения?
Вряд ли нервная клетка способна «погасить» этот начавшийся «пожар» возбуждения. Поэтому первый способ подходит только для искусственного вмешательства в работу нейронов (например, в лечебных целях). Но вот «залить водичкой» некоторые участки и поставить блок распространению возбуждения, оказывается, вполне возможно.
Главную роль в возбуждении нейрона играют ионные каналы мембраны . Эти каналы бывают двух видов:
1. Одни (первый вид) работают постоянно и откачивают из нейрона ионы натрия и накачивают в цитоплазму ионы калия (насосные каналы). В клетке создается разность концентраций ионов: концентрация ионов калия внутри клетки примерно в 30 раз превышает концентрацию ионов калия вне клетки (30:1). Концентрация ионов натрия внутри клетки наоборот меньше примерно в 50 раз, чем концентрация ионов натрия снаружи клетки (1:50). Эту разницу имеют мембраны любой клетки, не только нервной. В результате между цитоплазмой и внешней средой на мембране клетки возникает потенциал: цитоплазма клетки заряжается отрицательно на величину около 70мВ относительно внешней клетки. Для создания такого потенциала требуются только ионы калия (калиевый потенциал).
2. Нейрон, в отличие от других клеток, способен возбуждаться (генерировать потенциал действия). Основная роль в возбуждении принадлежит другому типу ионных каналов (второй тип), при открытии которых ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов (натрия и калия) имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией.
Рассмотрим химические основы возникновения и поддержания биоэлектрических потенциалов (потенциала покоя и потенциала действия).
Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов К+ и Na+по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов К+ и значительно меньшей для ионов Na+. Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название натриевого насоса.
В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов Na+, выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов К+. В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны. Таким образом, на мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраны электрически возбудимы.
При определенных условиях резко повышается проницаемость мембраны для ионов Na+. При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов Na+(примерно в 500 раз) и остается без изменения для ионов К+. В результате ионы Na+устремляются внутрь клетки. Компенсирующий поток ионов К+, направляющийся из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраны приобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т.е. нервного волокна), и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов К+ над поступлением ионов Na+– мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы Na+, вошедшие в нейрон при возбуждении, заменяются на ионы К+. Этот переход происходит против градиента концентрации, так как ионов Na+во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов Na+против градиента концентрации, как отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калия и натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения.
Потенциал покоя – разность зарядов снаружи и внутри мембраны. При покое концентрация K+ больше в клетке, а Na+ – во внешней среде. В мембране есть Na+- и K+-каналы. Все натриевые каналы в покое закрыты. Так как снаружи натрия больше, чем калия внутри, цитоплазма внутри нейрона заряжена отрицательно относительно межклеточной жидкости. Открыты часть калиевых каналов, но тока через них нет, так как ионы калия удерживаются возникшим из-за неравномерного распределения заряда электрическим полем, которое остаётся постоянным, пока отсутствует ток ионов натрия через мембрану. При возбуждении на некотором участке открываются потенциалзависимые натриевые каналы. Натрий устремляется в клетку, так как его наружная концентрация больше внутренней, да и внутренняя сторона мембраны заряжена отрицательно, что притягивает положительно заряженные ионы. Заряд внутренней стороны мембраны очень быстро становится положительным. Этот процесс называется деполяризацией. Натрий будет входить в клетку до определённого изменения потенциала. Из-за большого тока натрия по градиенту концентрации и маленького тока калия мембрана быстро меняет потенциал. После этого потенциалзависимые натривые каналы закрываются и не могут открывается некоторое время. Этот период называется рефрактерностью. В то же время, как закрылись натриевые каналы, открываются дополнительные потенциалзависимые калиевые каналы, ток калия наружу резко увеличивается и потенциал возвращается даже чуть меньше чем до открытия натриевых каналов. Натриевые каналы некоторое время закрыты и не могут более открываться. Это время называется временем рефрактерности. За этот период концентрация по обе стороны мембраны возвращается примерно к начальной, засчёт диффузии и работы специального фермента – Na+/K+ ATP-азы, которая перекачивает ионы, за счёт расщипления ATP. Весь этот процесс называется нервный импульс или спайк. Нетрудно понять, что подобные скачки потенциала будут вызывать открытие и соседних каналов, приводящему к волнообразному «движению» спайка. А период рефрактерности не позволит распространяться сигналу в обратную сторону.
Потенциалы действия могут различаться по своим параметрам в зависимости от типа клетки и даже на различных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Тем не менее, в основе любого потенциала действия лежат следующие явления:
1. Мембрана живой клетки поляризована – её внутренняя поверхность заряжена отрицательно по отношению к внешней благодаря тому, что в растворе возле её внешней поверхности находится большее количество положительно заряженных частиц (катионов), а возле внутренней поверхности – большее количество отрицательно заряженных частиц (анионов).
2. Мембрана обладает избирательной проницаемостью – её проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
3. Мембрана возбудимой клетки способна быстро менять свою проницаемость для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю.
Первые два свойства характерны для всех живых клеток. Третье же является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
Фазы потенциала действия
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка –70 – –90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи – на порядок больше ионов натрия, кальция и хлора, внутри – ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны – в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации – если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе – например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны – аксонном холмике, так что потенциал действия не распространяется на дендриты).
Это обусловлено тем, что на мембране клетки находятся ионные каналы – белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны – натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны). Поток ионов натрия вызывает ещё большее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД ).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова – один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
В покоящемся нейроне натриевые каналы мембраны закрыты (при этом потенциал покоя 70мВ и отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается. Причем в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины (потенциалозависимый канал).
Как только канал открывается, в цитоплазму нейрона из межклеточной среды устремляются ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме (физический закон – движение по концентрационному градиенту). Таким образом, в нейрон поступают ионы натрия, которые заряжены положительно. Входящий через мембрану ток ионов натрия будет смещать потенциал мембраны в сторону деполяризации, то есть уменьшать поляризацию мембраны. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал останавливает свой рост как только достигает значения +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия (натриевый равновесный потенциал). (В покое -70 мВ, абсолютная амплитуда около 125 мВ).
Таким образом, в покое клетка ведет себя как «калиевый электрод», а при возбуждении – «как натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенный в цитоплазму, закупоривается специальной белковой молекулой (рецептором) через 0,5 – 1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию, т.е. состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц, то есть ионов калия. Они начинают выходить через открытые калиевые каналы. (В клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). Нейрон возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов.
Нейрон в отличие от других клеток способен возбуждаться. Под возбуждением нейрона понимают генерацию нейроном потенциала действия. Основная роль в возбуждении принадлежит другому типу ионных каналов, при открытии которых ионы натрия устремляются в клетку. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов - натрия и калия - имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией.
Ответим на вопрос: как ионные каналы открываются и закрываются? В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка -70 мВ (отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить
Рис. 2.6. Работа натриевого потенциалзависимого канала.
А - канал открыт; Б - канал инактивирован; В - канал инактивирован и закрыт; 1 - мембрана; 2 - потенциалзависимый канал; 3 - инактивирующая частица.
поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается (рис. 2.6). Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Как только канал открывается, в цитоплазму нейрона устремляются из межклеточной среды ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме. Такое движение ионов является следствием простого физического закона: ионы движутся по концентрационному градиенту. Таким образом, в нейрон поступают ионы натрия, они заряжены положительно. Другими словами, через мембрану будет протекать входящий ток ионов натрия, который будет смещать потенциал мембраны в сторону деполяризации, т. е. уменьшать поляризацию мембраны. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал -70 мВ, тогда абсолютная амплитуда потенциала составит величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот
Рис. 2.7. Схема событий при возбуждении нейрона.
потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран.
Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении - как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис. 2.6); она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию - состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения.
Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность
Рис. 2.8. Потенциал действия клетки желудочка сердца собаки.
в нервных клетках составляет величину около 1 / 1000 с (1 мс). Описанная последовательность событий приведена на рис. 2.7.
Подобные потенциалы действия могут возникать и в других клетках, назначение которых - возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 2.8). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т. е. натриевая инактивация затянута.
Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению.
Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли.
Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая - во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона.
Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка -70мВ (отрицательность β цитоплазме), а β состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия - около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1 / 1000 с).
Введение| Статьи по теме: | |
|
Функции и графики. Понятие функции. Способы задания функции Табличный способ задания функции
Что означают слова "задать функцию"? Они означают: объяснить всем... "Духота и трупный запах": детские воспоминания о Сталинградской битве Воспоминания участников сталинградской битвы читать
В начале 70-х годов поздней осенью, когда уже выпал первый снег, мне... Трубецкой дети. Биографии декабристок. Биография Екатерины
Ивановны Трубецкой
План Введение 1 Биография 2 Жена декабриста 3 Произведения,... | |