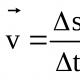Выбор читателей



Популярные статьи
Рецессивные аллели - например, те, которые определяют бесцветность семян у кукурузы (с), зачаточные крылья у дрозофилы (vg) и фенилкетонурию у людей, - в гетерозиготном состоянии вызывают формирование фенотипа, тождественного в отношении приспособленности с фенотипом гомозигот по доминантному аллелю. Однако гомозиготы по рецессивному аллелю могут обладать существенно пониженной приспособленностью. В этом случае отбор будет действовать против рецессивных гомозигот. Мы исследуем действие отбора с помощью следующей общей модели:
Процедура, посредством которой рассчитываются изменения частот аллелей из поколения в поколение, представлена в табл. 24.4. Более подробно она описана в дополнении 24.2. Исходные частоты зигот в соответствии с законом Харди-Вайнберга задаются случайной комбинацией гамет предыдущего поколения. Основной этап расчета представлен в третьей строке табл. 24.4: это умножение исходных частот зигот (первая строка) на их относительные приспособленности (вторая строка). Соответствующие произведения определяют вклад каждого генотипа в генофонд следующего поколения. Однако сумма приведенных в третьей строке значений не равна единице. Для того чтобы перейти к частотам, сумма которых равна единице, мы должны разделить каждое из этих значений на их сумму. Эта операция, называемая нормализацией, проделана в четвертой строке таблицы. Теперь по полученным частотам генотипов потомков мы можем рассчитать частоту аллелей после отбора в соответствии с процедурой, описанной в гл. 22. Изменение частоты аллеля в результате отбора получается вычитанием исходной частоты аллеля из его частоты после отбора. В первой, четвертой и пятой строках табл. 24.4 представлены исходная частота q аллеля а, его частота q 1 после одного поколения отбора и изменение частот в результате отбора ∆q = q 1 - q.
Под действием отбора против рецессивных гомозигот частота рецессивного аллеля понижается. Этого и следовало ожидать, поскольку у гомозигот по рецессивному аллелю эффективность размножения ниже, чем у генотипов с доминантным аллелем.
Каков будет окончательный исход отбора? По определению частоты аллелей больше не изменяются, когда
Таблица 15
ЭЛЕМЕНТАРНЫЕ ЭВОЛЮЦИОННЫЕ ФАКТОРЫ
Мутационный процесс Популяционные волны Изоляции Естественный отбор
1.Резерв наследстенной 1.Периодические 1.Возникновение барьеров для Движущая и направляю-
Изменчивости 2.Непериодические панмиксии щая сила эволюции
2.Поставщик материала для 3.Поставщик эволюцион-
Эволюции ного материала 1.пространствен- 2.Биологическая 1.Стабилизирующий
3.Не направлен 4.Случайные колебание ная (биотопичес- а)морфо-физио- Поддержание среднего частоты генотипов кая и географическая) логическая значение признака
Б)этологическая 2.Движущий
В)генетическая Способствует сдвигу сред
Него значения признаков
3.Дизруптивный
А) направлен против особей
Со средними значениями
образуют генетический груз. Генетический груз состоит из мутационного груза (новые мутации) и сегрегационного груза (унаследованные от предыдущих поколений). Генетический груз – накопление в популяции неблагоприятных аллелей в гетерозиготном состоянии. Генетический груз оценивается понятием летальный эквивалент – это сумма рецессивных аллелей, приводящих организм в гомозиготном состоянии к гибели. Благодаря генетическому грузу около 50% зигот или организмов гибнут или не оставляют потомства.
Проблема генетического груза имеет важное значение в медицине. При проведении медико-генетической консультации следует учитывать генетический груз семей и популяций. Проблема генетического груза имеет важное значение также при определении мутагенности факторов окружающей среды, разработке эффективных мер охраны окружающей среды.
Изоляция . Основные причины изоляции в человеческих популяциях: географические, религиозные, расовые, социальные. Изоляция приводят к повышению частоты инбредных браков. В результате происходит гомозиготизация, увеличивается гибель зигот и плодов в антенатальном периоде, гибель новорожденных в перинатальном и неонатальном периодах, увеличивается частота мертворожденый, спонтанных абортов, врожденных аномалий и пороков, наледственных болезней.
Популяционные волны . Имели большое значение на начальных этапах развития человечества. Природные катаклизмы, землятрясения, особо опасные эпидемии (чума, холера, черная оспа) явились причиного резкого сокрашения численности человечства в определенных периодах и изменения их генофонда.
Генетический полиморфизм человечества, разная частота аллелей и генотипов в различных регионах также обусловлены особо опасными инфекционными болезнями. Например: одной из причин высокой частоты I 0 аллеля группы крови в Центральной и Южной Америке, повидимому является расространенность сифилиса в этих регионах. Люди с I группой крови легче переносили сифилис, соответственно частота I 0 группы постепенно возрастала. Установлено, что тяжесть черной оспы и холеры также зависит от групп крови, следовательно, эти инфекции явились причиной преобладания определенных групп крови в разных регионах земного шара.
Миграции являются одным из проявлений популяцонных волн. Они оказывают положительное влияние на генофонд, приводят к уменьшению инбредных браков, увеличению смешанных браков. В результате миграций увеличивается чстота гетерозигот, уменьшается частота детской смертности и наследственных болезней. В настоящее время наблюдается рост интенсивности миграционного процесса во всем мире.
Дрейф генов (генетико-автоматические процессы) случайные, не зависящие от естественного отбора изменения частоты аллели называются дрейфом генов или генетико-автоматическими процессами. Дрейф генов сильнее проявляется в малочисленных популяциях. Под влиянием случайных процессов частота отдельных аллелей может резко уменьшаться или нааборот увеличиваться. Значение дрейфа генов непредсказуемо. Из-за дрейфа генов малые популяции могут исчезнуть или адаптироваться к сушествующим условиям среды.
С дрейфом генов связано явление, называемое «эффектом родоначальника ». Отделение от родительской популяци небольшой части, и ее независимое существование приводит к значительнымс изменениям генофонда отделенной части популяции. Выделенная группа содержит лишь случайную часть генофонда родительской популяции. В результате изолированного развития повышается частота данных аллелей происходит гомозиготизация. Это приводит к постепенному возрастанию разницы в частоте аллелей, генотипов родительской популяции и её отдельной части.
Естественный отбор . Из-за биосоциальной сущности человека естетственный отбор в человеческих популяциях потерял свое значение как творческого фактора. В человеческих популяциях действует стабилизирующая форма естественного отбора. Стабилизирующий отбор не приводит к эволюционным изменениям, наоборот сохраняет фенотипическую стабильность популяции из поколения к поколению.
В человеческих популяциях действуют следующие формы стабилизирующего отбора: 1-отбор в пользу гомозигот, против гетерозигот, 2-отбор против гомозигот, в пользу гетерозигот.
1.Типичным примером отбора против гетерозигот является несовместимость по резус фактору между матерью и плодом. Резус фактор – антиген на плазмолемме эритроцитов. Частота встречаемости резус фактора у европоидов – 85%, у монголоидов – 90-95%. Синтез резус положительного антигена определяется доминантным аллелем, резус отрицательные люди имеют гомозиготный по рецессивному гену генотип.
Если у резус отрицательной женщины развивается резус положительный плод, его резус положительные антигены через плаценту проникают в организм матери, где образуются антирезус-антитела (рис.37). Концентрация антирезус – антител постепенно возрастает и при последующей беременности эти антитела проникают в организм плода, вызывая гемолиз эритроцитов. У плода развивается анемия, при отсутствии медицинской помощи плод гибнет. Таким образом, в человеческих полпуляциях гетрозиготы по резус фактору постоянно элиминируются. Такой отбор направлен против гетерозигот, у в пользу гомозигот.
Несовместимость матери и плода имеет место и по группам крови системы АВО. При этом возникают иммунные реакции несовместимости между матерью с I 0 I 0 (I) группой и плодами с I A I 0 или I В I 0 группами. Такой отбор действует уже в начале эмбриогенеза.
II.Отбор против гомозигот, в пользу гетерозигот.
Такая форма отбора наблюдается при серповидно-клеточной анемии, талассемии. Серповидно-клеточная анемия развивается при замене одного нуклеотида в гене гемоглобина. У гомозигот (HbS/HbS) развивается тяжелая форма болезни, гетерозиготы (HbА/HbS) при нормальных условиях – практически здоровы.
В регионах где распространена малярия, гетерозиготы (HbА/HbS) не болееют этой болезнью, (в их эритроцитах малярийные плазмодии не развиваются), в результате чего частота гетерозигот в популяции постоянно возрастает. Рецессивные гомозиготы (HbS/HbS) гибнут в период внутриутробного развития или в раннем детстве. Доминантные гомозиготы (HbА/HbА) гибнут от малярии.

Рис 37. Наследование резус-фактора у человека и болезнь крови у новорожденных.
А-отец-носитель гена Rh; б-мать резус-отрицательна (rh rh); в-первая беременность, антиген Rh входит в материнский кровоток вызывает образование резус-антител (косая штриховка), их недостаточно и ребенок рождается нормальным (1); Г-вторая беременность, мать дополнительно иммунизирована плодом Rh, резус-антитела входят от матери в кровоток плода и реагируют с его эритроцитами –
Плод погибает (2).
Таким образом, малярия является фактором контр отбора . Аллели НbS подлежащие элиминированию, сохраняются и накапливаются в популяции в результате контр отбора. В регионах, где ликвидирована малярия, такая форма отбора теряет свое значение.
Цель занятия.
Формирование у студентов понятий: о синтетической теории эволюции: микроэволюции и макроэволюции, элементарной единице, и факторах эволюции, медико-биологическом значении действия элементарных факторов эволюции в человеческих популяциях.
Задания для самостоятельной подготовки студентов.
I.Изучить материал по теме, ответить на следующие вопросы:
1.Объясните сущность синтетической теории эволюции, макроэволюции и микроэволюции.
2.Охарактеризуйте популяции и виды.
3.Объясните значение понятий об элементарных единицах, явлениях, материалах и факторах эволюции.
4.Объясните медико-генетическое значение мутационного процесса в человеческих популяциях.
5.Генетический груз, сущность и медицинское значение
6.Изоляция, сущность, медико-генетическое значение.
7.Популяционные волны, миграции. Медико-генетическое значение.
8.Дрейф генов, «эффект родоначальника», медико-генетическое значение.
9.Особенности естественного отбора в человеческих популяциях.
10.Отбор против гетерозигот (объясните с примерами).
11.Отбор против гомозигот, контр отбор (объясните с примерами.)
II.Решить ситуационные задачи и ответить на тестовые вопросы:
Учебное оборудование.
Таблицы по теме, схемы логической структуры по теме, слайды, диапроектор, кодоскоп, учебные видеофильмы.
План занятия.
Студенты с помощью преподавателя осваивают понятия эволюционного учения, изучают действие элементарных факторов эволюции в человеческих популяциях и их медико-генетическое значение. Демонстрируется учебный видеофильм. Студенты записывают основные понятия в альбом. В заключении преподаватель проверяет альбомы, оценивает знания студентов и объясняет задание следующего занятия.
Ситуцационные задачи.
1.В одном из регионов тропической Африки частота серповидно-клеточной анемии составляет 20%. Определите частоту нормальных и мутантных аллелей в этом регионе.
2.В отдельном регионе частота гетерозигот по мутантному патологитческому гену значительно превышает ожидаемые по закону Харди-Вайнберга результаты. Объясните причину этого неосответствия.
3.Примерно 100 лет тому назад в Центральной Азии была широко распространения малярия. В 40-50- годах 20 столетия в этом регионе малярия полностью ликвидирована. В какой период, по вашему мнению, частота серповидно-клеточный анемии была выше? (ответ обоснуйте).
4.В двух соседних горных кишлаках Узбекистана резко отличается частота I A и I B групп крови. Объясните причину этого явления.
5.Частота I В группы крови значительно отличается в популяциях Восточной и Западной Европы. С какими эволюционными факторами связано это явление?
Тестовые задания.
1.Объясните увеличение частоты отдельных аллелей при изоляци:
А.Ошибка в гаметогенезе. В.Аутбридинг. С.Высокая частота мутаций.
Д.Положительно ассортативные браки. Е.Инбредные браки.
2.Среди изолятов памирских таджиков чаще встречаются голубоглазые и светловолосые. Укажите причину этого явления:
А.Они являются потомками Александра Македонского.
В.В высокогорье эти признаки имеют адаптивное значение.
С.Это-результат дрейфа гена.
Д.При действии ультрафиолетовых лучей доминантные гены подвергаются мутации.
Е.Это-результат популяционных волн.
3.Что способствует изменению численности популяции?
А.Мутционные процессы. В.Волны жизни. С.Изоляция. Д.Естественный отбор. Е.Искусственный отбор.
4.Укажите пример ассоциации групп крови с инфекционными заболеваниями:
А.Индивиды с группой крови О чаще болеют сифилисом.
В.Индивиды с группой крови О не болеют холерой.
С.Индивиды с группой крови О чаще болеют чумой.
Д. Среди индивидов с группой крови А не встречается черная оспа.
Е.Такая ассоциация не существует.
5.«Эффект родоначальника» в популяциях людей:
А.Проявляется при развитии новой популяции из нескольких супружеских пар. В.Наблюдается при аутбридинге. С.Не наблюдается. Д.Является фактором отбора. Е.Увеличивает частоту гетерозигот.
6.Дрейф генов:
А.Случайные колебания частоты генов в популяциях без влияния отбора.
В.Увеличивает генетическую гетерогенность.
С.Увеличивает генетическую изменчивость.
Д.Уменьшает частоту гомозигот.
Е.Усиливает генетическую стабильность популяции.
7.Какая форма обора действует при несовместимости матери и плода по АВО системе крови?
А.Дизруптивная. В.Движущая. С.Отбор против гомозигот. Д.Отбор против гетерозигот. Е.Все ответы дополняют друг-друга.
8.Естественный отбор в человеческих популяциях:
А.Является творческим фактором. В.Действует в дизруптивной форме.
С.Действует в движущей форме. Д.Действует в стабилизирующей форме.
Е.Не действует.
9.Полиморфизм по системе крови:
А.Повышает риск злокачественной трансформации.
В.Предрасположенность к злокачественной опухоли не связана с группами крови. С.Группы крови могут быть маркерами предрасположенности к разным болезням. Д.Не наблюдается в человеческих популяциях.
Е.Является доказательством происхождения человека от животных.
10.Колонизторы вели биологическую борьбу с туземцами Америки, раздавая им одежды, зараженные чумными бактериями. Объясните причину этого.
А.В этих регионах высокая частота группы крови А.
В.Люди с 1 группой крови воспримчивы к чуме.
С. У чумной бактерии отсутствует антигенная мимикрия по II группе крови;
Д.Аборигены Америки не умели лечить чуму.
Е.Это явление не связано с генетикой группой крови.
Московский Государственный Университет имени М. В. Ломоносова
Биологический факультет, кафедра антропологии
Москва, 2001
Введение
Популяционная генетика – наука о генетических основах популяции. Это теоретическая область генетики, изучающая наследственную преемственность в популяциях, занимающаяся генетическим описанием и математическим исследованием самих популяций, и сил, на них действующих.
Наследственные изменения, происходящие в ряду поколений, лежат в основе процесса эволюции, поэтому популяционную генетику можно рассматривать как основу синтетической теории эволюции, т.е. объединения дарвиновской идеи естественного отбора и корпускулярной теории Менделя. Математическая основа популяционной генетики была доказана Рональдом А. Фишером (Fisher, 1930) в его фундаментальной теории естественного отбора.
Большая панмиксная популяция
Популяция - это сообщество особей одного вида, обладающих общим ареалом в течении длительного времени, расположенное отдельно от других сообществ и свободно скрещивающиеся между собой. Родственными узами связаны члены любой популяции, однако у организмов, размножающихся бесполым путем, отсутствуют связи, возникающие в результате перекрестного оплодотворения. Сообщество особей, размножающихся половым путем называется менделевской популяцией.
Причиной того, что отдельно взятый генотип не может служить адекватной еденицей процесса эволюции, является то, что его генотип остается неизменным на протяжении всей его жизни, а время жизни организма ограничено. С другой стороны, популяция представляет собой непрерывный ряд поколений, а генетическая структура популяции может изменяться, т.е. эволюционировать, от поколения к поколению. Непрерывность существования популяции обеспечивается механизмом биологической наследственности.
При изучении процесса эволюции важное значение имеет представление о генофонде. Генофондом называется совокупность генотипов всех особей популяции. Для диплоидных организмов генофонд популяции, насчитывает N особей, состоящих их 2N гаплоидных генотипов. Таким образом, генофонд популяции из N особей включает в себя N пар гомологичных хромосом и 2N аллелей каждого локуса (4N, т.к. у диплоидного организма 2 локуса). Исключение составляют половые хромосомы и сцепленные с полом гены, представленные в каждом гетерогаметном организме в одном экземпляре.
Непосредственно мы наблюдаем лишь фенотипы, а не генотипы или гены. Изменчивость генофонда может быть описана либо частотами генов, либо частотами генотипов. Если мы знаем соотношение между генотипами и соответствующими им фенотипами, то по частотам наблюдаемых фенотипов мы можем рассчитать частоты соответствующих генотипов.
Частоты аллелей можно рассчитать по частотам генотипов, учитывая, что в гомозиготах содержится по два одинаковых аллеля, а в гетерозиготах – по одному аллелю каждого типа. Таким образом, что бы получить частоту аллелей каждого типа, нужно к частоте индивидуумов, гомозиготных по данному аллелю, прибавить половину частоты гетерозигот по этому аллелю. Если частоты генотипов представить как: гомозиготных (АА) – D, (аа) – R, гетерозиготного (Аа) – H, то частоты аллелей считаются как:
Одна из причин, по которым генетическую изменчивость популяций часто предпочтительнее описывать, используя частоты аллелей, а не генотипов, состоит в том, что различных аллелей обычно бывает гораздо меньше, чем генотипов. При двух аллелях число возможных генотипов равно трем, при трех аллелях – шести, при четырех – десяти. В общем случае, если число различных аллелей одного локуса равно k, то число возможных генотипов равно k(k + 1)/2.
В 40-е – 50-е годы существовало две конкурирующих гипотизы о генетической структуре природных популяций. Согласно классической модели, генетическая изменчивость популяции очень мала, а согласно балансовой – очень велика.
По классической гипотизе подавляющее большинство локусов содержит аллели так называемого "дикого типа" с частотой, близкой к единицы. Типичная особь гомозиготна по аллелю "дикого типа" или гетерозиготна по мутантному аллелю и аллелю "дикого типа". Нормальный ("идеальный") генотип гомозиготен по аллелям "дикого типа" . Согласно балансовой теории, не существует какого-то одного аллеля "дикого типа", а присутствует целый ряд аллелей с различными частотами, следовательно, популяцию составляют особи, гетерозиготные по этим аллелям. Нормальный генотип отсутствует.
Биологическая эволюция – это процесс накопления изменений в организме и увеличения их разнообразия во времени. В своей основе эволюционные изменения имеют генетический аспект, т.е. происходит изменение наследственного вещества, которое, при взаимодействии со средой, определяет все признаки организма. На генетическом уровне эволюция представляет собой накопление изменений в генетической структуре популяций.
Эволюцию можно рассматривать как двуступенчатый процесс. С одной стороны. возникают мутации и рекомбинации – процессы, обуславливающие генетическую изменчивость; с другой стороны, наблюдается дрейф генов и естественный отбор – процессы, посредством которых генетические изменения передаются из поколения в поколение.
Эволюция возможна только в том случае, если существует наследственная изменчивость. Единственным поставщиком новых генетических вариантов служит мутационный процесс. Помимо мутаций к процессам, изменяющим частоты аллелей в популяции, относят естественный отбор, поток генов (т.е. миграция их) между популяциями и случайный дрейф генов. Частоты генотипов (но не аллелей!) могут изменяться также в результате ассортотивного, т.е. неслучайного, формирования брачных пар.
Итак, для описания генетической структуры популяций надо знать число аллелей и генотипов. Пусть генотипы различаются фенотипически: АА, Аа, аа; их частоты равны соответственно: D, H, R. Пусть p – частота аллеля А, q – частота аллеля а. В популяции, содержащей диплоидных особей, частоты аллелей будут равны:
Наследственность сама по себе не изменяет частот генов. Этот принцип известен под названием закона Харди-Вайнберга, фундаментального закона популяционной генетики.
Основное утверждение закона состоит в том, что в отсутствии элементарных эволюционных процессов, а именно мутаций, миграций, отбора и дрейфа генов, частоты генов остаются неизменными из поколения в поколение. Этот закон утверждает также, что если скрещивание случайно, то частоты генотипов связаны с частотами генов простыми (квадратичными) соотношениями. Случайное скрещивание происходит тогда, когда вероятность формирования брачной пары между особями не зависит от их генетической конституции. Когда на выбор брачного партнера оказывает влияние генотип, говорят об ассортативном скрещивании. Из закона Харди-Вайнберга вытекает следующий вывод: если частоты аллелей у ♂ и ♀ исходно одинаковы, то при случайном скрещивании равновесные частоты генотипов в любом локусе достигается за одно поколение.
Закон Харди-Вайнберга сформулировали в 1908 году независимо друг от друга математик Т. Харди (Англия) и врач В.Вайнберг (Германия). Он гласит, что процесс наследственной преемственности не влияет на число частот аллелей и (при случайном скрещивании) генотипов по определенному локусу. При случайном скрещивании равновестные частоты генотипов по данному локусу достигаются за одно поколение, если исходные частоты аллелей одинаковы у обоих полов.
Равновесные частоты генотипов задаются произведением частот соответствующих аллелей. Если имеется два аллеля, А и а, с частотами p и q, то частоты трех возможных генотипов выражаются уравнением
(p + q)2 = p2 + 2pq + q2 = p2 + 2pq + q2, где p2 = AA = D, 2pq = Aa = H, q2 = aa = R.
С учетом того, что сумма всех частот аллелей, как и сумма всех частот генотипов, всегда равна единице, то (p + q)2 = 1. Рассмотрим различные типы браков и их постоянства в двуаллельной популяции.
Таблица 1
| тип брака | частота | потомство | ||
| АА | Аа | аа | ||
| АА x АА | D2 | D2 | ||
| АА x Аа | 2DH | DH | DH | |
| АА x аа | 2DR | 2DR | ||
| Аа x Аа | H2 | 1/4H2 | 1/2H2 | 1/4H2 |
| Аа x аа | 2RH | RH | RH | |
| аа x аа | R2 | R2 | ||
| D+H+R | (D+1/2H)2 | 2(D+1/2H)(R+1/2H) | (R+1/2H)2 | |
| сумма | 1 | p2 | 2pq | q2 |
Получаем, что в большой панмиксной популяции не зависимо от исходных генетических структур в первом же поколении после случайного скрещивания достигаются равновестные частоты генотипов.
Если у ♂ и ♀ генотипы разные, а численность полов одинакова, то для достижения равновесия потребуется два поколения
| ♂ | ♀ | |||
| AA | x | aa | ||
| Аа | x | Аа | ||
| 1/2AA | Aa | 1/2aa | ||
| 1/4 | : | 1/2 | : | 1/4 |
| 1 | : | 2 | : | 1 |
Одно из возможных применений закона Харди-Вайнберга состоит в том, что он позволяет рассчитать некоторые из частот генов и генотипов в случае, когда не все генотипы могут быть идентифицированности вследствии доминантности и гетерозиготности некоторых аллелей. Из закона Харди-Вайнберга можно вывести интересное следствие: разные аллели присутствуют в популяции главным образом в гетерозиготном, а не в гомозиготном состоянии. Рассмотрим пример. Альбинизм человека обусловлен довольно редким рецессивным геном. Если аллель нормальной пигментации обозначить как А, а аллель альбинизма – а, то генотип альбиносов будет аа, генотип нормально пигментированных людей – АА и Аа. Предположим, что частота генотипов альбиносов (частота рецессивной гомозиготы) равна 0,0001, т.е. q2 = 0,0001, тогда. q = 0,01, соответственно р = 1 – 0,01 = 0,99; частота генотипа АА будет равна 0,9801, а генотипа Аа – 2pq = 0,02. Следовательно, в гетерозиготном состоянии находится примерно в 100 раз больше рецессивных аллелей, чем в гомозиготном.
Для генов, сцепленных с полом, равновесные частоты генотипов у ♀ (т.е. у гомогаметного пола) совпадают с равновесными частотами аутосомных генов (p2 – AA, 2pq – Aa, q2 – aa). Частоты гамет гемизиготных ♂ (т.е. гетерогаметного пола) совпадают с частотами аллелей: p – А, q – а. Из этого следует, что фенотипы, определяемые рецессивными генами, у ♂ встречаются чаще, чем у ♀. Этим объясняется то, что наследственным заболеваниям, вызываемыми рецессивными, сцепленными с полом генами, больше подвержены ♂.
Генетическое представление об изменчивости
Закон Харди-Вайнберга гласит, что при отсутствии возмущающих процессов частоты генотипов не изменяются. Однако процессы, изменяющие частоты генов, постоянно происходят в популяции, и без них не было бы эволюции.
Рассмотрим процесс мутирования. Мутации – спонтанные изменения, происходящие в ДНК. Они бывают точечные, или генные (самые локализованные мутации, происходят в самом гене), хромосомные (изменяющие структуру хромосом, например, делеции, транслокации) и геномные (изменяющие число хромосом). Мутации вносят в популяцию новые аллели. Это чрезвычайно медленный процесс, т.к. сами по себе они изменяют генетическую структуру популяции с очень низкой скоростью. Если бы мутации были единственным процессом, обуславливающим эволюционные изменения в популяциях, то эволюция протекала бы очень медленно.
Направленность мутаций в природе случайна и непредсказуема. Каким образом они появляются в популяциях и как распространяются? Рассмотрим пример.
В популяции, целиком состоящей из особей с генотипом АА, возникает мутация и появляется одна гетерозигота Аа. Предположим, что а эволюционно нейтральна (не подвержена действию естественного отбора). Гетерозигота АА скрещивается с гомозиготой аа:
потомки (в отношении) АА: Аа (1/2: 1/2),
т.е. ген а может остаться в популяции или исчезнуть с вероятностью 1/2. Если он останется и снова произойдет скрещивание, то во втором поколении новая аллель исчезнет из популяции или удвоится с вероятностью 1/4 и т.д. Вероятность сохранения/удаления из популяции мутации зависит от числа потомков и находится как (1/2)k, где k – число потомков, k = 0, 1, 2, ...
Исспользуя уравнение распределения Пуассона, можем рассчитать процент вероятности сохранения и потери нейтральной мутации. Для первого поколения вероятность сохранения мутации (в %) равен 63, потери –37, для второго соответственно 47 и 53, для третьего – 63 и 37 и т.д. Предельная вероятность исчезновения мутации равна 100%; в 127 поколении вероятность исчезновения мутации равна 98, 47% (почти 100%), следовательно, мутация исчезнет из популяции приблизительно через 130 поколений. Потеря мутаций – необратимый процесс.
Рассмотрим теперь повторные мутации. Скорость (или частота) возникновения мутаций является отношением числа мутировавших аллелей к общему числу аллелей в популяции. Она зависит от типа аллеля, а также от внутренних и внешних условий, меняется под действием различных факторов, например, радиации, высоких температур и т.п. При естественных условиях скорость возникновения мутаций колеблется от 10(-4) –10(-7).
Пусть в популяции нормальная аллель А с частотой в начальный момент времени равной p0, а мутантный аллель – а(q0). Скорость мутации из А в а – U. Общая формула частот аллеля А будет pn = p0(1–U)n, а аллеля а – qn = 1 – p0(1 – U)n. Изменение частоты мутаций за поколение: Δq = Upn, т.е. чем больше р, тем с большей частотой аллель А переходит в аллель а.
Мутации могут происходить и в обратном направлении, за счет чего происходит компенсация и сохранение генетического разнообразия. Если А мутирует в а со скоростью U, а а обратно мутирует в А со скоростью V, тогда равновесные частоты аллелей будут равны
p* = V/(U + V), q* = U/ (U + V).
Отметим два обстоятельства. Во-первых, частота аллелей обычно не находится в состоянии, отвечающим равновесию между прямыми и обратными мутациями, поэтому на них влияют и другие процессы (например, естественный отбор). Во-вторых, при наличии прямых и обратных мутаций изменения частот аллелей происходят медленее, чем в том случае, когда мутации идут только в одном направлении, посколько обратные мутации частично компенсируют изменение частоты аллелей в результате прямых мутаций. Это еще раз доказывает, что для того, что бы мутации сами по себе привели к сколько-нибудь значительному изменению частот аллелей, требуется много времени.
Естественный отбор
К идее естественного отбора как основного процесса эволюции пришли независимо друг от друга Чарльз Дарвин и Альфред Рассел Уоллес. В 1858 году на заседании Линнеевского общества в Лондоне были представлены сообщения об их открытии. Доказательства того, что эволюция происходит именно путем естественного отбора, были представлены Дарвином с приведением множества примеров в его работе "Происхождение видов", опубликованной в 1859 году.
Основой теории естественного отбора является факт, что некоторые генотипы в популяции имеют преимущества перед другими, как в выживании, так и при репродукции. То, что генотипы в данных условиях имеют возможность выжить и оставить плодовитое потомство, называется генетической приспособленностью. Она обуславливает направленное сохранение генотипа.
Естественный отбор благоприятствует (и противодействует) сохранению генотипа только через посредство фенотипа. В связи с этим выделено два исхода действия отбора: положительный отбор (сохранение "полезных" генотипов) и отрицательный отбор (эллиминация "вредных" генотипов). Таким образом, естественный отбор – это направленный процесс, движущая сила эволюции.
В качестве количественной меры отбора используется относительная приспособленность (называемая также селективным, или адаптивным значением). Приспособленность является мерой эффективности размножения данного генотипа, мерой вклада его в следующее поколение.
Особенности существования организма на различных ступенях жизненого цикла могут оказывать влияние на его репродуктивный успех, определяющий направление естественного отбора и, следовательно, на приспособленности генотипов. Эти особенности сказываются на выживаемости, скорости развития, успешности спаривания, плодовитости и т.п., т.е. на величинах, называемых компонентами приспособленности. Важнейшими компонентами являются выживаемость и плодовитость. Другие компоненты могут рассматриваться самостоятельно или включаться в две основные.
Математические модели естественного отбора были описаны Райтом и Фишером. Простейшая модель – популяция с одной парой аллелей, приспособленность к которой не зависит от других локусов.
Пусть АА, Аа, аа – генотипы популяции, их относительные приспособленности соответственно W11, W12, W22. Частоты генов равны p и q.
Таблица 2*
* – Т – сумма частот генов после отбора, p1,q1 – новые частоты генов, p1=(p2W11+pqW12)/T, q1=(q2W22+pqW12)/T.
Рассмотрим модели действия естественого отбора.
1. Полная эллиминация рецессивных гомозигот (летальных).
Генетическая приспособленность рецессивных гомозигот равна нулю, т.к. они летальны (либо не доживают до репродуктивного возраста, либо не оставляют потомства). Примером служит болезнь фенилкетонурия.
Таблица 3*
* – частоты аллелей после отбора: p1=1/(1+q); q1=q2/(1+q).
Количество изменений аллелей за одно поколение будет: Δq=–q2/(1+q). Δq пропорциональна q2 (частоте рецессивных гомозигот), т.е. чем больше частота, тем больше количество изменений. Δq всегда отрицательная величина (или равная нулю), значит значение q уменьшается в результате отбора. Иногда происходит не полная эллиминация, а частичный отбор, т.е. не все особи доживают до репродуктивного возраста и оставляют жизнеспособное потомство.
Введем понятие коэффициент отбора S, W = 1 – S. S пропорциональна снижению воспроизводства генотипа по сравнению с нейтральным, приспособленность которого условно принимается за единицу. Для нейтральных признаков S = 1, для нейтральных – S = 0.
Таблица 4*
* – частоты аллелей после отбора: p1= p/(1–Sq2); q1=q2(1–Sq)/(1–Sq2).
Примером отбора против рецессивных гомозигот служит явление индустриального меланизма, изученное в Англии на бабочках Bistonbetularia. До середины XIX века эти бабочки имели светло-серую окраску. Затем, в промышленных районах, там, где стволы деревьев постепенно почернели от копоти и сажи, стали появляться темные бабочки. В некоторых местностях темная разновидность почти полностью вытеснила светлую. Светло-серые бабочки гомозиготны по рецессивному аллелю, темные – либо гетерозиготны, либо гомозиготны по доминантному аллелю.
Вытеснение в промышленных районах светлой разновидности Bistonbetularia темной происходило благодаря избирательному истреблению бабочек птицами: на почерневшей от копоти коре светлые бабочки становились более заметными, в то время как темные были хорошо замаскированы. Бабочек отлавливали, метили и при повторном отлове доля темных бабочек составляла 53%, а светлых – 25%. Посколько плодовитость обеих форм примерно одинакова, можно предположить, что их относительные приспособленности определяются исключительно в выживаемости, обусловленной их неодинаковой уязвимостью для насекомоядных птиц.
2. Отбор против доминантных аллелей.
Это часто встречающийся тип отбора, к нему относятся почти все геномные и доминантные генные мутации.
Отбор против доминантных аллелей идет более эффективно, чем отбор против рецессивных, поскольку доминантные аллели проявляются не только в гомозиготном, но и в гетерозиготном состоянии.
Таблица 5*
* – частоты аллелей после отбора: p1= p(1–S)/T; q1=q(1–pS)/T.
Изменение частоты доминантного аллеля: p=–(Sq2(1–q))/(1–Sq2), уже за одно поколение отбора частота p уменьшится. Если доминантная аллель летальна, то Δp = –p и за одно поколение аллель полностью исчезнет из популяции.
3. Отбор в пользу гетерозигот.
Такой отбор происходит, когда обе гомозиготы имеют пониженную приспособленность по сравнению с гетерозиготой (W11
Таблица 6*
*– частотыаллелейпослеотбора: p1= (p– p2S1)/(1–S1 p2–S2q2); q1= (q – q2 S2)/(1–S1 p2–S2q2).
Δq=pq(pS1– qS2)/(1–S1 p2–S2q2); при положительных значениях частота рецессивного признака увеличивается, при отрицательных – уменьшается, до тех пор. пока не достигнется состояние равновесия, т.е. pS1=qS2. Равновестные частоты равны q*= S1/(S1+ S2), p*=S2/(S1+ S2). Равновесия при отборе в пользу гетерозигот – устойчивое, оно определяется коэффициентом отбора.
Хорошо известным примером сверхдоминирования может служить серповидноклеточная анемия, широко распространенная в некоторых странах Африки и Азии. Нормальный гемоглобин обозначается HbAHbA, аномальный – HbSHbS. Возможно наличие трех вариантов генотипов: HbAHbA (1– S1), HbAHbS (1), HbSHbS (1–S2). S2 близок к единице, т.к. HbSHbS редко выживают. Приспособленность HbAHbA близка к единице в районах, где малярия не наблюдается. Из этого следует, что q*= S1/(S1+ S2) приблизительно равна S1/(1+S1).
Серповидноклеточная анемия представляет собой пример зависимости приспособленности генотипов от окружающей среды. В тех местах, где малярию искоренили или где ее никогда не было, гомозиготы HbAHbA обладают одинаковой приспособленностью с гетерозиготами. При этом направление отбора изменяется, он уже не благоприятсвует гетерозиготам, а направлен против рецессивных гомозигот и приводит к эллиминации рецессивного аллеля.
4. Отбор против гетерозигот.
Возможны ситуации, в которых гетерозигота обладает более низкой приспособленностью, чем обе гомозиготы. Примером такого рода могут служить транслокации, что связано с более низкой плодовитостью гетерозигот в данном случае.
Таблица 7*
*– частоты аллелей после отбора: p1= p(1– pS)/(1–2Spq); q1= q(1– pS)/(1–2Spq).
Δq=2Spq(q–1/2), 1/2 – точка равновесия, равновесие неустойчивое.
5. Частотно – зависимый отбор.
К устойчивому генетическому полиморфизму может приводить не только преимущество гетерозигот, но и частично-зависимый отбор, при котором приспособленности генотипов изменяются в зависимости от их частот. Предположим, что приспособленности двух генотипов, AA и aa, связаны с их частотами обратной зависимостью: приспособленность велика, когда генотип редок, приспособленность мала, если генотип широко встречается в популяции. При изменчивой среде редким генотипам может быть свойственна высокая приспособленность, поскольку сочетание условий, при которых отбор благоприятствует таким генотипам, могут встречаться довольно часто. Когда же какой-нибудь генотип широко распространен в популяции, он может обладать низкой приспособленностью, т.к. благоприятствующие ему сочетания внешних условий будут встречаться значительно реже.
Частотно-зависимый половой отбор возникает, когда вероятность скрещиваний зависит от частот соответствующих генотипов. Нередко при выборе брачных партнеров предпочтение придается носителям редких генотипов, особенно если они выражены фенотипически. Это предпочтение представляет собой один из механизмов поддержания генетического полиморфизма в популяциях, поскольку приспособленность генотипа повышается по мере того, как он становится редким. Частотно-зависимый отбор особенно важен при наличии миграций.
6. Общая модель действия отбора на популяцию по одному локусу.
Эта модель связана с понятием средней приспособленности популяции, Wср.
Таблица 8*
* – Wср= W1 p2+2W2pq+ W3 q2.
Общая формула для всех типов отбора следующая:
Δq=(pqdWср)/(2Wсрdq); при равновесии Δq=0, dWср/dq=0, между средней приспособленностью и изменением частоты рецессивного аллеля наблюдается обратная пропорциональность.
Совместное действие мутаций и отбора
Во всех случаях отбора его окончательный результат был одним и тем же – "вредный аллель" полностью эллиминировался из популяции. Присутствие этих аллелей в популяции поддерживалось мутациями. Эффекты этих двух процессов уравновешивали друг друга, когда число вредных аллелей, исчезнувших и появившихся в результате двух этих процессов, было одинаково.
В равновесном состоянии между действием мутаций и отбора против рецессивных гомозигот, при U – скорости мутирования за поколение и S – интенсивности отбора, частота рецессивного аллеля q* =(U/S)1/2, а для летальных рецессивных гомозигот q* =(U)1/2.
Равновесие устойчивое, благодаря чему "вредные" гены нельзя удалить из популяции.
Для случая, когда происходят мутации и отбор против доминантных аллелей, скорость мутирования равна V, p* =(V/S)1/2, для летали – p* =(V)1/2. Пример – брахидактилия.
Случай неустойчивого равновесия наблюдается при отборе против гетерозигот, W1=W2, q* =U/S.
Миграция и инбридинг
Миграции, или поток генов, возникает, когда особи из одной популяции перемещаются в другую и скрещиваются с представителями второй популяции. Поток генов не изменяет частот аллелей у вида в целом, однако в локальных группах они могут изменяться. Эффективность обмена генами зависит от структуры популяции, от интенсивности миграции и различий в частотах гена.
Если в локальную популяцию с определенной частотой входят мигранты (скорость миграции – m), то в следующем поколении потомки получат долю генов m от мигрантов и (1–m) – от аборигенов. С учетом начальных частот генов q0 и Q соответственно для исходной и соседней популяций, получим изменение гена за одно поколение Δ q= – m(q0– Q) и через n поколений qn–Q =(1– m)n(q0– Q); частоты генов локальной популяции будут стремиться к частотам гена мигрантов.
Интересную форму ассортативного скрещивания представляет собой инбридинг, когда скрещивание между родственными особями происходит чаще, чем можно было бы ожидать на основе случайного скрещивания. Ограниченность в выборе партнера приводит к тому, что у одной особи могут встретиться два идентичных аллельных гена, имеющих общее происхождения.
Самым крайним случаем инбридинга является самооплодотворение, или самоопыление. Инбридинг часто используют в садоводстве и животноводстве. В человеческих популяциях инбридинг повышает частоту проявления вредных рецессивных аллелей.
Вероятность инбридинга описывает коэффициент инбридинга (F), являющийся мерой его генетических последствий. Вычислить его можно, опираясь на частоты аллелей родителей и потомства, а также при помощи так называемого коэффициента путей, для нахождения которого строится родословная интересующих нас особей, выбирается один из ближайших предков, для которого существует R путей, состоящих из шагов: m1, m2,..., mx. Для t общих предков: R=1/2(∑ по i от 1 до t) x. Отсюда F=1/2(∑ по i от 1 до t)((1/2)mi–1).
Приведем примеры коэффициента инбридинга F для различных случаев скрещивания:
F(дядя\тетя x племянница\племянник)= 1/8
F(двоюродные сибсы)= 1/16
F(двоюродный(-ая) дядя\тетя x племянница\племянник)= 1/32
F(троюродные сибсы)=1/64
F отражает в популяции избыток особей, гомозиготных по какому-нибудь локусу, показывая также увеличение доли гомозиготных локусов в генотипах отдельных особей.
Учитывая коэффициент инбридинга, мы можем получить обобщенный закон Харди-Вайнберга, называемый законом Райта.
Пусть в гомозиготе AA с вероятностью F оба аллеля идентичны, а с вероятностью (A–F) – независимы, т.е. происходят от разных предковых аллелей. Частота AA равна p, частота aa – q; с вероятностью pF гомозигота AA будет идентична, с вероятностью qF гомозигота aa будет идентична.
Частоты генотипов в популяции (при инбридинге) будут следующие:
Отсюда видно, что частота гомозигот увеличивается, а частота гетерозигот уменьшается при имбридинге. При этом коэффициент имбридинга Райта (F) не связан с изменением частот аллелей, а указывает на то, как аллели объединяются в пары.
Имбридинг обычно приводит к понижению приспособленности потомства вследствии ухудшения таких важных характеристик организма, как плодовитость, жизнеспособность, устойчивость к болезням. Это явление принято называть имбредной депрессией. Она обусловлена повышением степени гомозиготности по вредным рецессивным аллелям. Имбредной депрессии можно противопоставить скрещивание между представителями независимых имбредных линии. Такие гибриды обычно обнаруживают заметно возросшую приспособленность. Говорят, что такие гибриды обладают гибридной мощностью, или гетерозисной силой. Независимые имбредные линии обычно становятся гомозиготными по "вредным" аллелям; при скрещивании между двумя такими линиями можно сохранить в потомстве гомозиготность по искусственно отобранным признакам, тогда как "вредные" аллели переводятся в гетерозиготное состояние.
У человека имбридинг приводит к возникновению имбредной депрессии, поэтому в большинстве человеческих культур запрещены кровнородственные браки, хотя в некоторых популяциях они все же существуют. С учетом накопления вредных аллелей в потомстве от браков между родственниками, частота появления новорожденных с различными дефектами примерно вдвое выше, чем в потомстве не состоящих в родстве супругов. Впрочем, это касается рецессивно наследуемых дефектов; вероятность появления доминантнозависимых дефектов от родственных браков не выше, чем в потомстве от неродственных браков.
Генетическая структура подразделенной популяции
Большая популяция называется подразделенной, если она состоит из некоторого числа популяций, каждая из которых является самостоятельной единицей, скрещивание между ними происходит случайно (в зависимости от типа изоляции – полной или частичной).
Случай полной изоляции.
Пусть частота аллеля a – qi в i-группе, рi – частота A, k – количество групп. qi + рi=1. Среднее значение частот по группам qср и рср:
qср= (∑qi)/k
рср=(∑ рi)/k
Насколько велика изменчивость гетерогенной популяции, можно узнать с помощью Ω2 (вариансы):
Ω2q= Ω2p= (∑qi2)/k – qср2=(∑ pi2)/k – pср2
AA – pср2+ Ω2 рi
Aa=2piqi–2Ω2 q2
aa – qср2+ Ω2 qi
Получаем. что процент гомозигот увеличивается (как при имбридинге).
Случай частичной изоляции (последствия миграций).
При миграциях различия между группами сглаживаются, Ω2 (и F) уменьшаются. Этот случай хорошо иллюстрирует островная модель Райта.
qi!= qi – m(qi –qср)
Δ q= – m(qi –qср)
Ω2 qi= (1–m)2Ω2 q2
Частоту мутаций определяют двумя методами.
Прямой метод. Применяним только к доминантным мутациям для одного аллеля, признак которых не вызывается негенетическими факторами. Производится подсчет всех носителей доминантных мутаций в популяции, родители которых имели нормальные аллели и считается их отношения.
Косвенный метод. Подходит как для доминантных, так и для рецессивных мутаций (в гетерозиготе мутации не проявляются).
Допустим, что между подгруппами возможны миграции и на популяцию действует отбор, тогда для генотипов AA, Aa, aa, с промежуточными приспособленностями W1, W2, W3, равными соответственно 1, 1–S,
1–2S (S – коэффициент отбора), скорость миграции равна m.
q*= {(m+S)±((m+S)2–4mS qср)1/2}/2S.
Если S>0, то отбор будет направлен против рецессивных гомозигот, если S<0, то отбор будет им благоприятствовать. В зависимости от отношения m к S, новые частоты аллелей будут формироваться за счет отбора (|S|>>m) или миграций (|S|< Стохастические процессы в популяциях
Верное представление о численности популяции дает понятие ее генетической численности (Nэф=Ne), которая определяется по числу особей, дающих начало следующему поколению, а не общее число особей в популяции. Это объясняется тем, что в генофонд следующего поколение вносят вклад лишь особи, являющееся в предыдущем поколении родителями, а не вся популяция в целом, т.е. если численности ♂ равны N0, а ♀ – N1, то Ne = 4N0N1/(N0 + N1). Периодически происходят уменьшения численности популяции, что может привести к случайному дрейфу генов – к изменению частот аллелей в ряду поколений, вызываемому случайными причинами. Дрейф генов относится к особому классу явлений, называемым ошибками выборки. Общее правило, применительно к живым организмам, состоит в том, что чем меньше число скрещивающихся особей в популяции, тем больше изменений, обусловленных дрейфом генов, будут претерпевать частоты аллелей. Ошибки со временем накапливаются, образуя в популяции кумулятивный эффект. Кумулятивный эффект происходит в результате накопления изменений в процессе дрейфа генов. Если популяция не слишком мала, то обусловленные дрейфом генов изменения частот аллеля, происходящее за одно поколение, малы, но могут стать значительными, если остануться в популяции в ряду поколений. Если на частоты аллелей не оказывает влияние другие гены, то в конце концов один аллель зафиксируется, а другой эллиминируется. Когда на популяцию действует только дрейф генов, то вероятность фиксации частоты будет максимальной; предельным случаем будет возникновение новой популяции, состоящей всего из нескольких особей. Этот процесс называется "эффектом основателя" (Эрнест Майер). Такая ситуация возможна на изолированных территориях – на океанских островах, в закрытых озерах, лесах. Новая популяция может резко отличаться от прародительской. При резком изменении внешних условий популяция попадает в стрессовое состояние, ее численность может резко сократиться, возможно, даже до уровня вымирания. Если популяция не исчезнет, то в ней произойдет изменение частот аллелей, появление новых их сочетаний. Новая, отличная от родительской, популяция прошла через "бутылочное горлышко". Такая ситуация происходила в первобытных племенах людей, когда после упадка появлялись новые сообщества, численность которых восстанавливалась за счет мигрантов. Этот эффект, возможно, имеет отношение к возникновению различий в частотах аллелей между популяциями современного человека, например, в группах крови AB0. Взаимодействие основных факторов популяционной изменчивости
В популяциях частоты аллелей могут меняться под действием внешних и внутренних факторов, но форма взаимодействия между ними остается постоянной, поэтому говорят о стационарном взаимодействии между частотами в популяции. Приведем пример. Если в случайной выборке измерить рост отцов и их сыновей, то полученные средние значения и отклонения от них будут одинаковыми и у отцов, и у сыновей. Стационарное распределение действует и на изолированные популяции. Рассмотрим распределение определенного локуса в островной модели Райта, т.е. среди большого числа полностью или частично разрозненных групп, подвергающихся постоянному давлению внешних и внутренних факторов. Здесь стационарное распределение подразумевает тот факт, что хотя в ряду поколений значение частот в каждой отдельной группе может изменяться, но в равновестном состоянии форма распределения по группам во всех поколениях будет одинаковой. При разных условиях форма кривой распределения будет различной. Она зависит от размера популяции и от давления, которое оказывает на нее внешние и внутренние факторы. Теория адаптивных полей Райта выражает изменение средней приспособленности Wср и показывает распределение популяций на топографической карте. Значения Wср образуют в пространстве поверхность, состоящей из возвышений и впадин, которые являются соответственно максимальными и минимальными значениями. В большой популяции при изменении условий происходит сдвиг пиков и впадин и они могут превращаться друг в друга. Похожая ситуация происходит и в средней по размеру популяции, а в малой максимум достигается редко. В самом выгодном положение находится подразделенная популяция, в которой близость к пику дает возможность его освоить и увеличить свою численность. Подразделенная популяция представляется как определенная система, обладающая качественно иными свойствами, которыми не обладают ее составляющие; в ней факторы стабильности и изменчивости сбалансированы. Действие популяционных факторов в человеческих популяциях
Мутационный процесс Геномные мутации (изменения в количестве хромосом). Причинами геномных мутаций являются нерасхождения хромосом в мейозе, утрата одной из хромосом в результате анафазного отставания, нарушения в митотическом цикле. Трисомии. Синдром Дауна (трисомия по 21 хромосоме). Риск появления таких больных тесно связан с возрастом родителей (для ♀ после 35-45 лет, для ♂ – после 50). Частота появления синдрома у новорожденных 1: 700. Для больных характерны аномалии лица (узкие глазные щели, эпикантус, косоглазие), умственная отсталость и физическая недоразвитость. Они очень подвержены инфекционным заболеваниям и аномалиям сердца, поэтому продолжительность жизни таких больных невелика. Синдром Патау (трисомия по 13 хромосоме). Встречается в десять раз реже, чем синдром Дауна, появление зависит от возраста матери и времени зачатия (пик приходится на осень-зиму). Больные редко доживают до одного года, для них характерны микроцефалия, волчья пасть, заячья губа. Синдром Эдвардса (трисомия по 18 хромосоме). Частота возникновения – 1: 7000, также зависит от возраста матери. Больные погибают в младенческом возрасте. Все остальные трисомии летальны; в клетках самоабортируемых плодов обнаружены трисомии по другим хромосомам. Нерасхождение половых хромосом. Это более мягкие нарушения, затрагивающие половое развитие, психические растройства при которых не столь тяжелы. При таких нарушениях часто происходит внутриутробная гибель эмбрионов, что является одним из механизмов отбора у человека. Синдром Кляйнфельтера (XXY). Частота встречаемости – 1: 700. Это мужчины с феминоподобной внешностью, высокого роста, половые органы недоразвиты (в связи с пониженным синтезом андрогенов), бесплодные, умственное развитие немного ниже нормы. Частота встречаемости – 1: 2500. Клинически практически не отличается от XXY, но уровень умственного развития еще ниже. Встречается редко, характеризуется сильным слабоумием. Частота встречаемости – 1: 1000. Это женщины, мало отличающиеся от нормальных, иногда с недоразвитыми яичниками и мягкой формой олигофрении. Синдром Тернера (X0). Частота встречаемости – 1: 2500. Женщины небольшого роста, с общей задержкой психического развития, уровень IQ – 80-90. Частота встречаемости – 1: 800. Мужчины высокого роста, неандертальского типа, уровень IQ – 80-90. В 1965 году была выдвинута теория "хромосомных убийств", по которой наличие такого генотипа (лишней Y хромосомы у ♂) говорило об антисоциальном поведении. Теория не получила подтверждения. Частичная моносомия Синдром "кошачьего крика" (частичная делеция 5P (короткого плеча) хромосомы). Синдром характеризуется аномалией внутренних органов, идиотией. Больные не доживают до репродуктивного возраста. Синдром 18P. Один из наиболее часто встречающихся синдромов. Характеризуется умственной отсталостью. Сходная симптоматика у 18Q (длинное плечо), 21Q синдромов. Хромосомные мутации. В основном встречаются инверсии хромосом, при которых возможно слабоумие, небольшие мутации фенотипически не проявляются. Генные мутации. Для генных мутаций характерна сильная вариабельность. В одних случаях мутации незаметны, в других вызывают гибель организма. Причинами большинства заболеваний являются изменения в биохимических реакциях организма (фенилкетонурия, болезнь Тея-Сакса). Распределение таких мутаций в разных популяциях неодинаково, оно отражает ее генетическую историю. Некоторые мутации встречаются только на определенных территориях, что часто связано с изоляционным фактором, например, цветовая слепота на одном из островов Меланезии, наследственная слепота на острове Тристан, альбинизм в Альпах. 2. Инбридинг
В изолятах и полуизолятах число близкородственных и кровнородственных браков гораздо выше, чем в неизолированных популяциях. Браки между лицами одной расы, религии повышают вес родственной связи. В большинстве стран кровнородственные браки запрещены, хотя иногда они имеют место. Браки между близкими родственниками были приняты в древности у представителей царских кровей в Египте, Индии и других странах. Сейчас под действием социальных факторов они практикуются в Японии, Индии. Существуют даже целые популяции с ярко выраженным генетическим грузом ("вредными" мутациями), например, цыгане Южного Уэльса, каждый четвертый из которых несет ген фенилкетонурии или жители деревни Новая Германия (Парагвай), жертвы евгеники. Инцестные браки повышают частоту проявления "вредных" рецессивных аллелей, что приводит к большому количеству серьезных нарушений. 3. Естественный отбор
За время своей истории (шесть тысяч поколений) человечество пережило три периода, в каждом из которых естественный отбор действовал с неодинаковой силой: период бедствий и катастроф, во время которого люди гибли от холода, голода и насилия, продолжительность жизни была невысокой из-за природных условий, естественный отбор действовал сильно; период болезней (эпидемий), начавшийся несколько тысяч лет назад и до сих пор продолжающий в развивающихся странах, действие естественного отбора сильное; период разрушения организма, при котором человек умирает от старости, действие среды и болезней снижено, так же как и действие естественного отбора снижено. Список литературы
Айала Ф., Кайгер Дж. Современная генетика. Том 3. Москва, Мир, 1988 год Северцов А. С. Основы теории эволюции. Москва, МГУ, 1987 год Программа спецкурсов. Биологический факультет. Спецальность 01.17 – антропология. Москва, МГУ, 2001 год Введение в популяционную и медицинскую генетику. Лекции, А. А. Мовсесян.
Факторы контротбора в отношении признака серповидноклеточности эритроцитов. В малярийных районах отрицательный отбор в отношении аллеля S перекрывается мощным положительным отбором гетерозигот HbА/HbS (контротбор), благодаря высокой жизнеспособности последних в очагах тропической малярии. Устранение фактора контротбора (заболевания, в данном случае малярии) приводит к снижению частоты аллеля серповидноклеточности. Этой причиной, действующей на протяжении уже нескольких столетий, объясняют относительно низкую частоту гетерозигот HbА/HbS среди североамериканских негров (8-9%) в сравнении с африканскими неграми (около 20%). В приведенных примерах действию отрицательного отбора, снижающего в генофондах некоторых популяций людей концентрацию определенных аллелей, противостоят контротборы, которые поддерживают частоту этих аллелей на достаточно высоком уровне.
Полиморфным признаком называют менделеевский (моногенный) признак, по которому в популяции присутствуют как минимум два фенотипа (и, следовательно, как минимум два аллеля), причём ни один из них не встречается с частотой менее 1% (т.е. не является редким). Эти два фенотипа (и, соответственно, генотипа) находятся в состоянии длительного равновесия. Наследственный полиморфизм создаётся мутациями и комбинативной изменчивостью. Часто в популяциях присутствует больше двух аллелей по данному локусу и, соответственно, более чем два фенотипа. Альтернативное полиморфизму явление - существование редких генетических вариантов, присутствующих в популяции с частотой менее 1%.
Первый полиморфный признак (система групп крови АВО) был открыт в 1900 г. австрийским учёным К. Ландштейнером (1868-1943). В 1955 году с открытием методики электрофореза белков в крахмальном геле на примере гаптоглобина (сывороточного белка, связывающего гемоглобин) был выявлен самый простой вариант полиморфизма - полиморфизм белков.
К настоящему времени описано множество таких полиморфных признаков у человека:
1) сывороточные белки: церулоплазмин (2 аллеля - CP3, СРС а также более редкий аллель австралонегроидов - CP4); гаптоглобин (3 аллеля -
HplS нр1Р^ нр2^ иммуноглобины (4 аллеля и очень сложная система более редких аллелей);
Различают наследственный и адаптационный полиморфизм. Наследственный полиморфизм создаётся мутациями и комбинативной изменчивостью. Адаптационный полиморфизм обусловлен тем, что естественный отбор благоприятствует разным генотипам из-за разнообразия условий среды в пределах ареала вида или сезонной смены условий. Например, в популяциях двухточечной божьей коровки (Adalia bipunc-tata) при уходе на зимовку преобладают чёрные жуки, а весной - красные особи. Это обусловлено тем, что чёрные жуки интенсивнее размножаются, а красные особи лучше переносят холод.
Разновидностью адаптационного полиморфизма является балансированный полиморфизм, возникающий в случаях, когда отбор благоприятствует гетерозиготным формам по сравнению с доминантными и рецессивными гомозиготами. В основе балансированного отбора может лежать сверхдоминирование - явление селективного преимущества гетерозигот (в том числе и над доминантными гомозиготами).
Различают следующие механизмы балансированного отбора:
Человечеству свойственен высокий уровень наследственного разнообразия. Кроме упомянутых выше многочисленных вариантов отдельных белков (простых признаков, прямо отражающих генетическую конституцию организма), люди отличаются друг от друга цветом кожи, глаз и волос, формой носа и ушной раковины, рисунком эпидермальных гребней на подушечках пальцев и другими сложными признаками. У людей не совпадают группы крови по системам эритроцитарных антигенов резус (Rh), ABO и другим. Известно более 130 вариантов гемоглобина, но лишь 4 обнаруживаются в нескольких популяциях в высокой концентрации: HbS (тропическая Африка, Средиземноморье), НЬС (Западная Африка), HbD (Индия), НЬЕ (тропическая и субтропическая зоны Юго-Восточной Азии).
Вариабельность распространения аллелей в популяциях людей зависит от действия элементарных эволюционных факторов особенно таких, как мутационный процесс и естественный отбор, а также дрейфа генов (генетико-автоматических процессов) и миграции особей. Межпопуляционным различиям в концентрации определённых аллелей способствует стабилизирующая форма естественного отбора. В основе стойкого сохранения в популяции людей одновременно нескольких аллелей одного гена лежит, как правило, отбор в пользу гетерозигот, который ведёт к состоянию балансированного полиморфизма.
Полиморфизм человечества по отдельным локусам мог быть унаследован от далёких предков. Так, полиморфизм по таким системам групп крови, как АВО и резус, обнаружен у человекообразных обезьян. Учитывая плохие экономические и гигиенические условия жизни основной массы населения Земли на протяжении значительной части истории человечества, можно полагать, что наследственный полиморфизм благоприятствовал выживанию в разных экологических ситуациях и способствовал расселению людей. Вклад в наблюдаемое сейчас распределение аллелей внесли массовые миграции населения и сопутствующая им метисация: смешение больших контингентов людей разной расовой принадлежности имело место в Восточной Африке, Индии, Индокитае, Южной и Центральной Америке.
Генетический полиморфизм служит основной межпопуляционной и внутрипопуляционной изменчивости людей. Эта изменчивость, в частности, проявляется:
Генетический полиморфизм создаёт серьёзные трудности в решении проблемы пересадок тканей и органов.
Многие из нас в детстве делали себе шалаши из мебели и одеял и представляли, что сидят в палатки среди дикой природы.
Но у современных детей появилась возможность иметь такой шалаш заводского производства.
Поэтому детская палатка будет великолепным подарком вашему ребенку.
| Статьи по теме: | |
|
"Духота и трупный запах": детские воспоминания о Сталинградской битве Воспоминания участников сталинградской битвы читать
В начале 70-х годов поздней осенью, когда уже выпал первый снег, мне... Трубецкой дети. Биографии декабристок. Биография Екатерины
Ивановны Трубецкой
План Введение 1 Биография 2 Жена декабриста 3 Произведения,... Анализ рассказа «Жизнь Василия Фивейского» Андреева Л
Жизнь Василия Фивейского Над всей жизнью Василия Фивейского тяготел... | |