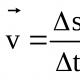Выбор читателей



Популярные статьи
Эндоплазматическую сеть (ЭПС), или эндоплазматический ретикулум (ЭПР) , удалось обнаружить только с появлением электронного микроскопа. ЭПС есть только в эукариотических клетках и представляет собой сложную систему мембран, образующих уплощенные полости и трубочки. Все вместе это выглядит как сеть. ЭПС относится к одномембранным органоидам клетки.
Мембраны ЭПС отходят от внешней мембраны ядра и по строению сходны с ней.
Эндоплазматическая сеть делится на гладкую (агранулярную) и шероховатую (гранулярную). Последняя усеяна прикрепленными к ней рибосомами (из-за этого и возникает «шероховатость»). Основная функция обоих типов связана с синтезом и транспортом веществ. Только шероховатая отвечает за белок, а гладкая - за углеводы и жиры.

По своему строению ЭПС представляет собой множество парных параллельных мембран, пронизывающих почти всю цитоплазму. Пара мембран образует пластинку (полость внутри имеет разную ширину и высоту), однако гладкая эндоплазматическая сеть в большей степени имеет трубчатое строение. Такие уплощенные мембранные мешочки называют цистернами ЭПС .
Рибосомы, расположенные на шероховатой ЭПС, синтезируют белки, которые поступают в каналы ЭПС , созревают (приобретают третичную структуру) там и транспортируются. У таких белков сначала синтезируется сигнальная последовательность (состоящая преимущественно из неполярных аминокислот), конфигурация которой соответствует специфическому рецептору ЭПС. В результате рибосома и эндоплазматическая сеть связываются. При этом рецептор образует канал для перехода синтезируемого белка в цистерны ЭПС.
После того, как белок оказывается в канале эндоплазматического ретикулума сигнальная последовательность от него отделяется. После этого он свертывается в свою третичную структуру. При транспортировке по ЭПС белок приобретает ряд других изменений (фосфорилирование, образование связи с углеводом, т. е. превращение в гликопротеин).
Большинство белков, оказавшихся в шероховатой ЭПС, далее попадают в аппарат (комплекс) Гольджи. Оттуда белки либо секретируются из клетки, либо поступают в другие органоиды (обычно лизосомы), либо откладываются как гранулы запасных веществ.
Следует иметь в виду, что не все белки клетки синтезируются на шероховатой ЭПС. Часть (обычно меньшая) синтезируется свободными рибосомами в гиалоплазме, такие белки используются самой клеткой. У них сигнальная последовательность не синтезируется за ненужностью.
Основной функцией гладкой эндоплазматической сети является синтез липидов (жиров). Например, ЭПС эпителия кишечника синтезирует их из жирных кислот и глицерола, всасывающихся из кишечника. Затем липиды попадают в комплекс Гольджи. Кроме клеток кишечника, гладкая ЭПС хорошо развита в клетках, секретирующих стероидные гормоны (стероиды относятся к липидам). Например, в клетках надпочечников, интерстициальных клетках семенников.
Синтез и транспорт белков, жиров и углеводов не единственные функции ЭПС. В печение эндоплазматический ретикулум участвует в процессах детоксикации. Особая форма гладкой ЭПС - саркоплазматический ретикулум – присутствует в мышечных клетках и обеспечивает сокращение за счет перекачки ионов кальция.
Структура, объем и функциональность эндоплазматической сети клетки не является постоянной на протяжении клеточного цикла, а подвержены тем или иным изменениям.
На гранулярной ЭПС находятся рибосомы, гладкая и промежуточная лишены их. Гранулярный ЭР в основном представлен цистернами , а гладкий и промежуточный - в основном каналами . Мембраны цистерн, каналов и пузырьков могут переходить друг в друга. ЭР содержит полужидкий матрикс, характеризующийся особым химическим составом.
Главная функция гранулярного (шероховатого) эндоплазматического ретикулума – синтез белков.
Гранулярный ЭПР представлен системой плоских цистерн. На их мембране со стороны цитозоля расположены рибосомы, объединенные в полисомы. На рибосомах гранулярного ЭПР происходит синтез белков, которые в зависимости от их конечного назначения могут быть разделены на три группы:
В ЭПР происходят начальные этапы сортировки синтезированных белков. Растворимые белки первых двух групп целиком поступают в цистерны ЭПР, который обеспечивает их обособления от цитозоля. Мембранные белки после синтеза остаются в составе мембраны ЭПР.
Сборка любого белка начинается на свободных рибосомах в цитозоле. В ЭПР поступают только те пептиды, у которых первым синтезируется специфический гидрофобный сигнальный пептид. Особая частица, распознающая сигнал (SRP) связывается с сигнальным пептидом, временно блокирует синтез белка и затем направляет рибосому к мембране эндоплазматического ретикулума, где SRP присоединяется к своему рецептору. Доставленная таким образом к ЭПР рибосома прикрепляется своей большой субъединицей к специальному белку-рецептору, который участвует в образовании канала. Частица, выполнившая свою задачу, покидает рибосому. Прекращается ее блокирующее действие, и синтез белка возобновляется.
Растущая белковая цепь поступает в ЭПР через канал в мембране. Пока белок в виде петли перетаскивается в полость ЭПР, его гидрофобный сигнальный пептид остается погруженным в мембрану. При синтезе растворимых белков сигнальный пептид отрезается, и белок высвобождается в полость ЭПР. Трансмембранные белки остаются заякоренными в билипидном слое с помощью неотрезанного сигнального пептида или за счет другого гидрофобного участка-сигнала окончания переноса (стоп-пептида). При чередовании в полипептиде сигналов начала переноса и окончания переноса белок будет пронизывать билипидный слой несколько раз.
Синтезированные в шероховатом ЭПР белки подвергаются обработке. Наряду с отрезанием сигнального пептида, важнейшим преобразованием является гликозилирование (соединение с олигосахаридом). Здесь же происходят начальные этапы преобразования олигосахаридов в гликопротеинах. В цистернах гранулярного ЭПР обеспечивается также и правильное сворачивание синтезированных белковых молекул (гидрофобные участки ориентированы внутрь). Это препятствует образованию агрегатов, выпадающих в осадок. В гранулярном ЭПР происходит сборка липопротеиновых мембран. Здесь синтезируются не только мембранные белки, но и липиды мембран.
Ферментативный синтез фосфолипидов происходит на обращенной к цитозолю стороне мембраны. Специальные белки-транслокаторы фосфолипидов могут переносить часть липидов во внутренний слой, создавая тем самым асимметрию билипидного слоя. ЭПР поставляет с помощью транспортных пузырьков мембранные белки и липиды аппарату Гольджи, который в свою очередь с помощью транспортных пузырьков снабжает ими плазмалемму и мембраны лизосом.
К органеллам данной группы относятся , рибосомы, комплекс Гольджи, лизосомы, пероксисомы. Они осуществляют синтез органических соединений, их транспорт в процессе химической доработки из одного участка канальцевой сети в другой, накопление, перемещение, упаковку и экзоцитоз готовых продуктов синтеза.
Эндоплазматическая сеть и рибосомы . Эндоплазматическая сеть представлена канальцами и цистернами, которые анастомозируют и формируют в гиалоплазме трехмерную сеть. В состав сети входят гранулярные (содержащие на внешних поверхностях мембран рибосомы) и агранулярные (без рибосом) участки.
Рибосомы синтезируют все разнообразие клеточных белков. На светооптическом уровне рибосомы неразличимы, об их количестве в клетке можно судить по интенсивности окраски цитоплазмы общегистологическими (базофилия) или специальными гистохимическими реактивами и флюорохромами, маркирующими РНК. На субмикроскопическом уровне рибосомы выглядят как осмиофильные черные точки (диаметром около 20-25 нм), а их рабочие комплексы - полисомы - как группы, или розетки, осмиофильных точек.
Компоненты рибосом создаются в разных участках клетки: рибосомальные РНК синтезируются в ядрышке; рибосомальные белки - в цитоплазме. Последние поступают в ядро, где комплексируются с молекулами РНК и объединяются в рибосомальные субъединицы. Затем субъединицы РНК транспортируются из ядра через поры и находятся в цитоплазме либо в диссоциированном (неактивном), либо ассоциированном друг с другом (активном) состоянии. Работающие органеллы состоят из двух ассоциированных (малой и большой) субъединиц, которые удерживаются в обратимо связанном состоянии с помощью катионов магния. Большую субъединицу рибосом образуют разные молекулы РНК, имеющие сложную вторичную и третичную структуру, в комплексе с рибосомальными протеинами. Большая субъединица значительно крупнее малой и имеет форму полушара. Малая субъединица выглядит в виде маленькой шапочки. При ассоциации субъединиц в рибосому происходит закономерное взаимодействие их поверхностей.
Между субъединицами работающей рибосомы имеет место строгое "разделение труда" - малая субъединица ответственна за связывание информационной РНК, большая - ведает образованием полипептидной цепи. В клетке нефункционирующие рибосомы находятся в диссоциированном состоянии, в связи с чем получают возможность постоянно обмениваться субъединицами и постоянно обновляться. В рабочем режиме рибосомы (от 3 до 20-30 в группе) образуют стабильный комплекс - полисому, в котором они связаны нитью информационной РНК.
О степени развития в клетке можно судить по базофилии цитоплазмы, обусловленной присутствием большого количества рибосом; агранулярные участки эндоплазматической сети на светооптическом уровне не обнаруживаются. В большинстве клеток преобладает гранулярная сеть, и оба вида сети имеют диффузную организацию - их элементы располагаются в гиалоплазме свободно, без какой-либо упорядоченности. Синтез белка в гранулярной сети происходит на рибосомах и полисомах, а ее каналы и цистерны являются вместилищем и транспортными магистралями для перемещения белка в коплекс Гольджи для доработки.
Ширина и количество канальцев и цистерн сети в клетках варьируют в зависимости от их функционального состояния - при повышении функциональных нагрузок на клетку канальцы и цистерны сети становятся множественными и значительно расширяются. Канальцы эндоплазматической сети непосредственно связаны с перинуклеарным пространством клетки.
Значение гранулярной эндоплазматической сети состоит в синтезе мембранных белков и белков, предназначенных "на экспорт" и необходимых другим клеткам, либо используемых во внеклеточных физиологических реакциях. Этот вид сети присутствует во всех клетках организма человека (кроме зрелых спермиев), однако наиболее развит в тех клетках, которые специализированы на синтезе больших количеств белковых молекул. Таких видов клеток в организме человека сравнительно немного. Примером являются плазмоциты, синтезирующие антитела (или иммуноглобулины); клетки поджелудочной железы, вырабатывающие комплекс белковых пищеварительных ферментов (панкреатический сок); гепатоциты, синтезирующие широкий спектр белков плазмы крови, свертывающей и противосвертывающей систем, а также некоторые другие клетки. В этих клетках канальцы сети располагаются упорядоченно (в некоторых случаях - строго параллельно) в виде так называемой эргастоплазмы.
В малодифференцированных и неспециализированных клетках гранулярная эндоплазматическая сеть , как правило, слабо развита, в структуре клеток преобладают свободные поли- и рибосомы, обеспечивающие синтез белков, необходимых клетке для роста и дифференцировки.
Агранулярная эндоплазматическая сеть имеет вид коротких канальцев и пузырьков (везикул), которые диффузно располагаются по всей гиалоштзме.В большинстве клеток элементы агранулярной сети, как правило, немногочисленны. В клетках, вырабатывающих стероидные гормоны (клетки надпочечников, половых желез), агранулярная сеть хорошо развита и ее многочисленные пузырьки занимают большие площади, либо образуют муфты вокруг липидных включений - предшественников стероидных гормонов. В мембранах сети находятся ферменты стероидогенеза.
Помимо стероидогенеза , она участвует в синтезе и метаболизме липидов, полисахаров, триглицеридов, процессе детоксикации продуктов метаболизма лекарственных препаратов и эндогенных клеточных ядов. В канальцах агранулярной сети депонируются большие запасы катионов кальция.
Строение эндоплазматической сети
Определение 1
Эндоплазматическая сеть (ЭПС, эндоплазматический ретикулум) – сложная ультрамикроскопическая, очень разветвлённая, взаимосвязанная система мембран, которая более или менее равномерно пронизывает массу цитоплазмы всех эукариотических клеток.
ЭПС – мембранная органелла, состоящая из плоских мембранных мешочков – цистерн, каналов и трубочек. Благодаря такому строению эндоплазматическая сеть значительно увеличивает площадь внутренней поверхности клетки и делит клетку на секции. Внутри она заполнена матриксом (умеренно плотный рыхлый материал (продукт синтеза)). Содержание различных химических веществ в секциях неодинаково, потому в клетке как одновременно, так и в определённой последовательности могут происходить различные химические реакции в незначительном объёме клетки. Эндоплазматическая сеть открывается в перинуклеарное пространство (полость между двумя мембранами кариолемы).
Мембрана эндоплазматической сети состоит из белков и липидов (в основном фосфолипидов), а так же ферментов: аденозинтрифосфатазы и ферментов синтеза мембранных липидов.
Различают два вида эндоплазматической сети:
Замечание 1
Иногда выделяют ещё переходящую, или транзиторную (тЭС) эндоплазматическую сеть, которая находится в участке перехода одной разновидности ЭС в другую.
Гранулярная ЭС свойственна всем клеткам (кроме сперматозоидов), но степень её развития разная и зависит от специализации клетки.
Сильно развита грЭС эпителиальных железистых клеток (поджелудочной железы, вырабатывающих пищеварительные ферменты, печени – синтезирующих альбумины сыворотки крови), фибробластов (клеток соединительной ткани, продуцирующих белок коллаген), плазматических клеток (продуцирование иммуноглобулинов).
Агранулярная ЭС преобладает в клетках надпочечников (синтез стероидных гормонов), в клетках мышц (обмен кальция), в клетках фундальных желез желудка (выделение ионов хлора).
Другим видом мембран ЭПС являются разветвлённые мембранные трубочки, содержащие внутри большое количество специфических ферментов, и везикулы – маленькие, окружённые мембраной пузырьки, в основном находящиеся рядом с трубочками и цистернами. Они обеспечивают перенесение тех веществ, которые синтезируются.
Эндоплазматическая сеть – это аппарат синтеза и, частично, транспорта веществ цитоплазмы, благодаря которому клетка выполняет сложные функции.
Замечание 2
Функции обоих типов ЭПС связаны с синтезом и транспортом веществ. Эндоплазматическая сеть является универсальной транспортной системой.
Гладкая и шероховатая эндоплазматические сети своими мембранами и содержимым (матриксом) выполняют общие функции:
Кроме того, каждой из разновидностей сети свойственны свои специфические функции.
Агранулярная эндоплазматическая сеть, кроме названных функций, общих для обоих видов ЭС, выполняет ещё и свойственные только для неё функции:
детоксикационная функция – обезвреживание єкзогенных и эндогенных токсинов;
Пример 1
В почечных клетках (гепатоцитах) содержатся ферменты оксидазы, способные разрушать фенобарбитал.
ферменты органеллы берут участие в синтезе гликогена (в клетках печени).
Для гранулярной эндоплазматической сети, кроме перечисленных общих функций, свойственны ещё и специальные:
Функции зернистой эндоплазматической сети связаны с транспортом белков, которые синтезируются в рибосомах и расположены на её поверхности. Синтезированные белки поступают внутрь ЭПС, скручиваются и приобретают третичную структуру.
Белок, который транспортируется к цистернам, значительно изменяется на своём пути. Он может, например, фосфорилироваться или превращаться в гликопротеид. Обычный путь для белка – это путь через зернистую ЭПС в аппарат Гольджи, откуда он или выходит наружу клетки, или поступает к другим органеллам той же клетки, например, к лизосомам), или откладывается в виде запасных гранул.
В клетках печени как зернистая, так и незернистая эндоплазматическая сетка берут участие в процессах детоксикации ядовитых веществ, которые потом выводятся из клетки.
Как и внешняя плазматическая мембрана, эндоплазматическая сетка имеет избирательную проницаемость, вследствие чего концентрация веществ внутри и снаружи каналов сетки неодинакова. Это имеет значение для функции клетки.
Пример 2
В эндоплазматической сетке мышечных клеток больше ионов кальция, чем в её цитоплазме. Выходя из каналов эндоплазматической сетки, ионы кальция запускают процесс сокращения мышечных волокон.
Липидные компоненты мембран эндоплазматической сети синтезируются ферментами самой сети, белковый – поступает из рибосом, расположенных на её мембранах. В гладкой (агранулярной) эндоплазматической сети нет собственных факторов синтеза белка, потому считается, что эта органелла образуется в результате потери рибосом гранулярной эндоплазматической сетью.
| Статьи по теме: | |
|
"Духота и трупный запах": детские воспоминания о Сталинградской битве Воспоминания участников сталинградской битвы читать
В начале 70-х годов поздней осенью, когда уже выпал первый снег, мне... Трубецкой дети. Биографии декабристок. Биография Екатерины
Ивановны Трубецкой
План Введение 1 Биография 2 Жена декабриста 3 Произведения,... Анализ рассказа «Жизнь Василия Фивейского» Андреева Л
Жизнь Василия Фивейского Над всей жизнью Василия Фивейского тяготел... | |