Выбор читателей




Популярные статьи
Л инейные полипептидные цепи индивидуальных белков за счёт взаимодействия функциональных групп аминокислот приобретают определённую пространственную трёхмерную структуру, называемую "конформация". Все молекулы индивидуальных белков (т.е. имеющих одинаковую первичную структуру) образуют в растворе одинаковую конформацию. Следовательно, вся информация, необходимая для формирования пространственных структур, находится в первичной структуре белков.
В белках различают 2 основных типа конформации полипептидных цепей: вторичную и третичную структуры.
2. Вторичная структура белков - пространственная структура, образующаяся в результате взаимодействия между функциональными группами пептидного остова.
При этом пептидные цепи могут приобретать регулярные структуры двух типов: α-спирали
β-структрура Под β-структурой понимают фигуру, подобную листу, сложенному «гармошкой». Фигура формируется за счет образования множества водородных связей между атомами пептидных групп линейных областей одной полипептидной цепи, делающей изгибы, или между разными полипептидными группами.


Связи - водородные, они стабилизируют отдельные фрагменты макромолекул.
3. Третичная структура белков - трёхмерная пространственная структура, образующаяся за счёт взаимодействий между радикалами аминокислот, которые могут располагаться на значительном расстоянии друг от друга в полипептидной цепи.
Структурно состоит из элементов вторичной структуры, стабилизированных различными типами взаимодействий, в которых гидрофобные взаимодействия играют важнейшую роль
стабилизации третичной структуры белка принимают участие:
· ковалентные связи (между двумя остатками цистеина - дисульфидные мостики);
· ионные связи между противоположно заряженными боковыми группами аминокислотных остатков;
· гидрофильно-гидрофобные взаимодействия. При взаимодействии с окружающими молекулами воды белковая молекула «стремится» свернуться так, чтобы неполярные боковые группы аминокислот оказались изолированы от водного раствора; на поверхности молекулы оказываются полярные гидрофильные боковые группы.
4. Четвертичной структурой называют взаимное расположение нескольких полипептидных цепей в составе единого белкового комплекса. Белковые молекулы, входящие в состав белка с четвертичной структурой, образуются на рибосомах по отдельности и лишь после окончания синтеза образуют общую надмолекулярную структуру. В состав белка с четвертичной структурой могут входить как идентичные, так и различающиеся полипептидные цепочки. В стабилизации четвертичной структуры принимают участие те же типы взаимодействий, что и в стабилизации третичной . Надмолекулярные белковые комплексы могут состоять из десятков молекул.
Роль.
Образование пептидов в организме происходит в течение нескольких минут, химический же синтез в условиях лаборатории - достаточно длительный процесс, который может занимать несколько дней, а разработка технологии синтеза – несколько лет. Однако, несмотря на это, существуют довольно весомые аргументы в пользу проведения работ по синтезу аналогов природных пептидов. Во-первых, путём химической модификации пептидов возможно подтвердить гипотезу первичной структуры. Аминокислотные последовательности некоторых гормонов стали известны именно благодаря синтезу их аналогов в лаборатории.
Во-вторых, синтетические пептиды позволяют подробнее изучить связь между структурой аминокислотной последовательности и её активностью. Для выяснения связи между конкретной структурой пептида и его биологической активностью была проведена огромная работа по синтезу не одной тысячи аналогов. В результате удалось выяснить, что замена лишь одной аминокислоты в структуре пептида способна в несколько раз увеличить его биологическую активность или изменить её направленность. А изменение длины аминокислотной последовательности помогает определить расположение активных центров пептида и участка рецепторного взаимодействия.
В-третьих, благодаря модификации исходной аминокислотной последовательности, появилась возможность получать фармакологические препараты. Создание аналогов природных пептидов позволяет выявить более «эффективные» конфигурации молекул, которые усиливают биологическое действие или делают его более продолжительным.
В-четвёртых, химический синтез пептидов экономически выгоден. Большинство терапевтических препаратов стоили бы в десятки раз больше, если бы были сделаны на основе природного продукта.
Зачастую активные пептиды в природе обнаруживаются лишь в нанограммовых количествах. Плюс к этому, методы очистки и выделения пептидов из природных источников не могут полностью разделить искомую аминокислотную последовательность с пептидами противоположного или же иного действия. А в случае специфических пептидов, синтезируемых организмом человека, получить их возможно лишь путём синтеза в лабораторных условиях.
57. Классификация белков: простые и сложные, глобулярные и фибриллярные, мономерные и олигомерные. Функции белков в организме .
Классификация по типу строения
По общему типу строения белки можно разбить на три группы:
1. Фибриллярные белки - образуют полимеры, их структура обычно высокорегулярна и поддерживается, в основном, взаимодействиями между разными цепями. Они образуют микрофиламенты, микротрубочки, фибриллы, поддерживают структуру клеток и тканей. К фибриллярным белкам относятся кератин и коллаген.
2. Глобулярные белки - водорастворимы, общая форма молекулы более или менее сферическая.
3. Мембранные белки - имеют пересекающие клеточную мембрану домены, но части их выступают из мембраны в межклеточное окружение и цитоплазму клетки. Мембранные белки выполняют функцию рецепторов, то есть осуществляют передачу сигналов, а также обеспечивают трансмембранный транспорт различных веществ. Белки-транспортёры специфичны, каждый из них пропускает через мембрану только определённые молекулы или определённый тип сигнала.
Простые белки , Сложные белки
Помимо пептидных цепей, в состав многих белков входят и неаминокислотные группы, и по этому критерию белки делят на две большие группы - простые и сложные белки (протеиды). Простые белки состоят только из полипептидных цепей, сложные белки содержат также неаминокислотные, или простетические, группы.
Простые.
Среди глобулярных белков можно выделить:
1. альбумины - растворимы в воде в широком интервале рН (от 4 до 8,5), осаждаются 70-100%-ным раствором сульфата аммония;
2. полифункциональные глобулины с большей молекулярной массой, труднее растворимы в воде, растворимы в солевых растворах, часто содержат углеводную часть;
3. гистоны - низкомолекулярные белки с высоким содержанием в молекуле остатков аргинина и лизина, что обусловливает их основные свойства;
4. протамины отличаются еще более высоким содержанием аргинина (до 85 %), как и гистоны, образуют устойчивые ассоциаты с нуклеиновыми кислотами, выступают как регуляторные и репрессорные белки - составная часть нуклеопротеинов;
5. проламины характеризуются высоким содержанием глутаминовой кислоты (30-45 %) и пролина (до 15 %), нерастворимы в воде, растворяются в 50-90 % этаноле;
6. глутелины содержат около 45 % глутаминовой кислоты, как и проламины, чаще содержатся в белках злаков.
Фибриллярные белки характеризуются волокнистой структурой, практически нерастворимы в воде и солевых растворах. Полипептидные цепи в молекулах расположены параллельно одна другой. Участвуют в образовании структурных элементов соединительной ткани (коллагены, кератины, эластины).
Сло́жные белки́
(протеиды , холопротеины ) - двухкомпонентные белки, в которых помимо пептидных цепей (простого белка) содержится компонент неаминокислотной природы - простетическая группа. При гидролизе сложных белков, кроме аминокислот, освобождается небелковая часть или продукты её распада.
В качестве простетической группы могут выступать различные органические (липиды, углеводы) и неорганические (металлы) вещества.
В зависимости от химической природы простетических групп среди сложных белков выделяют следующие классы :
· Гликопротеиды, содержащие в качестве простетической группы ковалентно связанные углеводные остатки и их подкласс - протеогликаны, с мукополисахаридными простетическими группами. В образовании связи с углеводными остатками обычно участвуют гидроксильные группы серина или треонина. Большая часть внеклеточных белков, в частности, иммуноглобулины - гликопротеиды. В протеогликанах углеводная часть составляет ~95 %, они являются основным компонентом межклеточного матрикса.
· Липопротеиды, содержащие в качестве простетической части нековалентно связанные липиды. Липопротеиды, образованные белками-аполипопротеинами связывающимися с ними липидами и выполняют функцию транспорта липидов.
· Металлопротеиды, содержащие негемовые координационно связанные ионы металлов. Среди металлопротеидов есть белки, выполняющие депонирующие и транспортные функции (например, железосодержащие ферритин и трансферрин) и ферменты (например, цинксодержащая карбоангидраза и различные супероксиддисмутазы, содержащие в качестве активных центров ионы меди, марганца, железа и других металлов)
· Нуклеопротеиды, содержащие нековалентно связанные ДНК или РНК, в частности, хроматин, из которого состоят хромосомы, является нуклеопротеидом .
· Фосфопротеиды, содержащие в качестве простетической группы ковалентно связанные остатки фосфорной кислоты. В образовании сложноэфирной связи с фосфатом участвуют гидроксильные группы серина или треонина, фосфопротеинами являются, в частности, казеин молока :
· Хромопротеиды - собирательное название сложных белков с окрашенными простетическими группами различной химической природы. К ним относится множество белков с металлсодержащейпорфириновой простетической группой, выполняющие разнообразные функции - гемопротеины (белки, содержащие в качестве простетической группы гем - гемоглобин, цитохромы и др.), хлорофиллы;флавопротеиды с флавиновой группой, и др.
2. Защитная функция
3. Регуляторная функция
4. Сигнальная функция
5. Транспортная функция
6. Запасная (резервная) функция
7. Рецепторная функция
8. Моторная (двигательная) функция
Пептидные цепи белков организованы во вторичную структуру, стабилизированную водородными связями. Атом кислорода каждой пептидной группы образует при этом водородную связь с NH-группой, соответствующей пептидной связи. При этом формируются следующие структуры: а-спираль, структура и р-изгиб.
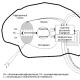
а-Спираль. Одной из наиболее термодинамически выгодных структур является правая а-спираль. На рис. 3.1 изображена а-спираль, представляющая устойчивую структуру, в которой каждая карбонильная группа образует водородную связь с четвертой по ходу цепи NH-группой. В а-спирали на один ее виток приходится 3,6 аминокислотного остатка, шаг спирали составляет примерно 0,54 нм, а расстояние между остатками - 0,15 нм. В а-спиральных участках торсионные углы ф и у равны 60 и 45 е и последовательно расположенные полипептидные звенья взаимно ориентированы.
L-Аминокислоты могут образовывать только правые а-спирали, причем боковые радикалы расположены по обе стороны оси и обращены наружу. В а-спирали полностью использована возможность образования водородных связей, поэтому она не способна в отличие от p-структуры образовывать водородные связи с другими элементами вторичной структуры. При образовании а-спирали боковые цепи аминокислот могут сближаться, образуя гидрофобные или гидрофильные компактные сайты. Эти сайты играют существенную роль при образовании трехмерной конформации белковой макромолекулы, так как используются для упаковки а-спиралей в пространственной структуре белка.
Рис. 3.1. а-Спираль белка аполипопротсина С-1 (но В. М. Степанову): а - гидрофильная поверхность: б - гидрофобная поверхность а-спирали белка
Спираль-клубок. Содержание а-спиралсй в белках неодинаково и является индивидуальной особенностью каждой белковой макромолекулы. Для некоторых белков, например для миоглобина, а-спираль лежит в основе структуры, другие, например химотрипсин, не имеют а-спирализованных участков. В среднем глобулярные белки имеют степень спирализации порядка 60-70%. Спирализованные участки чередуются с хаотическими клубками, причем в результате денатурации переходы спираль-клубок увеличиваются. Спирализация полипептидной цепи зависит от аминокислотных остатков, ее образующих. Так, отрицательно заряженные группы глутаминовой кислоты, расположенные в непосредственной близости друг от друга, испытывают сильное взаимное отталкивание, что препятствует образованию соответствующих водородных связей в а-спирали. По той же причине спирализация цепи затруднена в результате отталкивания близко расположенных положительно заряженных химических группировок лизина или аргинина. Большие размеры радикалов аминокислот также являются причиной, по которой спирализация полипептидной цепи затруднена (серин, треонин, лейцин). Наиболее часто интерферирующим фактором при образовании а-спирали является аминокислота пролин. Как известно, в пролине атом азота входит в состав жесткого кольца, что препятствует вращению вокруг связи N-С а. Кроме того, пролин не образует внутрицепочечную водородную связь из-за отсутствия при атоме азота водородного атома. Таким образом, во всех случаях, когда в полипептидной цепи встречается пролин, а-спиральная структура нарушается и образуется клубок или р-изгиб.
Р-Структура. В отличие от а-спирали p-структура образована за счет межцепочечных водородных связей между соседними участками полипептидной цепи, так как внугрицепочечные контакты отсутствуют. Если эти участки направлены в одну сторону, то такая структура называется параллельной (ср = -119°, ц/ = +113°) (рис. 3.2), если же в противоположную (у = +135°), то анти параллельной (рис. 3.3).

Рис. 3.2. Параллельная p-структура флаволокеина (по В. М. Степанову): пунктиром показаны водородные связи

Рис. 3.3.
Полипептидная цепь в p-структуре сильно вытянута и имеет не спиральную, а скорее зигзагообразную форму. Расстояние между соседними аминокислотными остатками по оси составляет 0,35 нм, т. е. в три раза больше, чем в а-спирали, число остатков на виток равно 2.
В случае параллельного расположения p-структуры водородные связи менее прочны по сравнению с таковыми при антипараллельном расположении аминокислотных остатков. В отличие от а-спирали, насыщенной водородными связями, каждый участок полипептидной цепи в p-структуре открыт для образования дополнительных водородных связей. Сказанное относится как к параллельной, так и к антипараллельной p-структуре, однако в антипарал- лельной структуре связи более стабильны. В отрезке полипептидной цепи, образующей p-структуру, находится от трех до семи аминокислотных остатков, а сама p-структура состоит из 2-6 цепей, хотя их число может быть и большим. p-Структура имеет складчатую форму, зависящую от соответствующих а-угле- родных атомов. Поверхность ее может быть плоской и левозакрученной таким образом, чтобы угол между отдельными отрезками цепи составлял 20-25° (рис. 3.4).

Рис. 3.4.

Рис. 3.5.
р-Изгиб. Глобулярные белки имеют шарообразную форму во многом благодаря тому, что для полипептидной цепи характерно наличие петель, зигзагов, шпилек, причем направление цепи может изменяться даже на 180°. В последнем случае имеет место p-изгиб (рис. 3.5).
Этот изгиб по форме напоминает шпильку для волос и стабилизируется одной водородной связью. Фактором, препятствующим его образованию, могут быть большие боковые радикалы, и поэтому довольно часто наблюдается включение в него наименьшего аминокислотного остатка - глицина. Эта конфигурация оказывается всегда на поверхности белковой глобулы, в связи с чем р-изгиб принимает участие во взаимодействии с другими полипептидными цепями.
Супервторичные структуры. Впервые супервторичные структуры белков были постулированы и затем обнаружены Л. Полингом и Р. Кори. В качестве примера можно привести суперспирализованную а-спираль, в которой две а- спирали скручены в левую суперспираль (рис. 3.6). Однако чаше суперспиральные структуры включают в себя как а-спирали, так и р-складчатыс листы. Их состав может быть представлен следующим образом: (cm), (ар), (ра) и (РХР). Последний вариант представляет собой два параллельных складчатых листа, между которыми находится статистический клубок (рСр), а-спираль (раР) или p-структура (РРР).
Соотношение между вторичной и супервторичной структурами имеет высокую степень вариабильности и зависит от индивидуальных особенностей той или иной белковой макромолекулы.
Домены - более сложные уровни организации вторичной структуры. Они представляют собой обособленные глобулярные участки, соединенные друг с другом короткими так называемыми шарнирными участками полипептидной цепи. Д. Бирктофт одним из первых описал доменную организацию химотрипсина, отметив наличие двух доменов у этого белка. Каждый из них имеет цилиндрическую форму, образованную p-структурой, и состоит из 6 антипараллельных цепей. В один из этих доменов входят 139 аминокислот с УУ-конца, другой - С-концевой включает в себя 115 аминокислотных остатков.

Рис. 3.6.
цилиндрами обозначены а-спирали; затемненные области - нсспирялизованные участки; стрелки - р-складчатые слои
Доменная организация характерна для многих белков. В этих белках, как правило, находится несколько структурных доменов, каждый из которых содержит до 200 аминокислотных остатков. Примером тому может быть белок глицеральдегидфосфатдегидрогеназа (ГАФД) (рис. 3.7).
В некоторых белках, например в иммуноглобулинах или сериновых про- тсиназах, структурные домены сходны по своей первичной структуре, что указывает на возможный механизм дубликации соответствующих генов, в других белках, например в гемоглобине, имеются определенные различия

Рис. 3.8. Домены гемоглобина человека: цилиндры - u-спирали; связывающие их нити - аморфные участки (по PDB-2001) (Yang, J., Kloek, А. Р., Goldberg, D. Е., Mathews, F. S.: Proc. Natl. Acad. Sci. USA, 92. p. 4224, 1995)

Рис. 3.7. Домены ГАФД из мышц омара (по А. А. Анисимову): а - НАД"-связывающий домен; 6 - каталитический домен
(рис. 3.8). По строению домены в белках разделяют на несколько групп в зависимости от содержания в них а-спиралей и ^-складчатых листов.
Таким образом, можно отметить следующее.
Строение белков. Структуры белков
Структуры белков: первичная, вторичная, третичная и четвертичная
Название «белки» происходит от способности многих из них при нагревании становиться белыми. Название «протеины» происходит от греческого слова «первый», что указывает на их важное значение в организме. Чем выше уровень организации живых существ, тем разнообразнее состав белков.
Белки образуются из аминокислот, которые соединяются между собой ковалентной – пептидной связью: между карбоксильной группой одной аминокислоты и аминогруппой другой. При взаимодействии двух аминокислот образуется дипептид (из остатков двух аминокислот, от греч. пептос – сваренный). Замена, исключение или перестановка аминокислот в полипептидной цепи вызывает возникновение новых белков. Например, при замене лишь одной аминокислоты (глутамина на валин) возникает тяжелая болезнь – серповидно-клеточная анемия, когда эритроциты имеют другую форму и не могут выполнять свои основные функции (перенос кислорода). При образовании пептидной связи отщепляется молекула воды. В зависимости от количества аминокислотных остатков выделяют:
– олигопептиды (ди-, три-, тетрапептиды и т. п.) – содержат до 20 аминокислотных остатков;
– полипептиды – от 20 до 50 аминокислотных остатков;
– белки – свыше 50, иногда тысячи аминокислотных остатков
По физико-химическим свойствам различают белки гидрофильные и гидрофобные.
Существуют четыре уровня организации белковой молекулы – равноценные пространственные структуры (конфигурации , конформации ) белков: первичная, вторичная, третичная и четвертичная.
Первичная структура белков является простейшей. Имеет вид полипептидной цепи, где аминокислоты связаны между собой прочной пептидной связью. Определяется качественным и количественным составом аминокислот и их последовательностью.
Вторичная структура образована преимущественно водородными связями, которые образовались между атомами водорода NH-группы одного завитка спирали и кислорода СО-группы другого и направлены вдоль спирали или между параллельными складками молекулы белка. Белковая молекула частично или целиком скручена в α-спираль или образует β-складчатую структуру. Например, белки кератина образуют α-спираль. Они входят в состав копыт, рогов, волос, перьев, ногтей, когтей. β-складчатую имеют белки, которые входят в состав шелка. Извне спирали остаются аминокислотные радикалы (R-группы). Водородные связи значительно более слабые, чем ковалентные, но при значительном их количестве образуют довольно прочную структуру.
Функционирование в виде закрученной спирали характерно для некоторых фибриллярных белков – миозин, актин, фибриноген, коллаген и т. п.
Третичная структура белка. Эта структура постоянна и своеобразна для каждого белка. Она определяется размером, полярностью R-групп, формой и последовательностью аминокислотных остатков. Полипептидная спираль закручивается и укладывается определенным образом. Формирование третичной структуры белка приводит к образованию особой конфигурации белка – глобулы (от лат. globulus – шарик). Его образование обуславливается разными типами нековалентных взаимодействий: гидрофобные, водородные, ионные. Между остатками аминокислоты цистеина возникают дисульфидные мостики.
Гидрофобные связи – это слабые связи между неполярными боковыми цепями, которые возникают в результате взаимного отталкивания молекул растворителя. При этом белок скручивается так, что гидрофобные боковые цепи погружены вглубь молекулы и защищают ее от взаимодействия с водой, а снаружи расположены боковые гидрофильные цепи.
Третичную структуру имеет большинство белков – глобулины, альбумины и т. п.
Четвертичная структура белка. Образуется в результате объединения отдельных полипептидных цепей. В совокупности они составляют функциональную единицу. Типы связей разные: гидрофобные, водородные, электростатические, ионные.
Электростатические связи возникают между электроотрицательными и электроположительными радикалами аминокислотных остатков.
Для одних белков характерно глобулярное размещение субъединиц – это глобулярные белки. Глобулярные белки легко растворяются в воде или растворах солей. К глобулярным белкам принадлежит свыше 1000 известных ферментов. К глобулярным белкам относятся некоторые гормоны, антитела, транспортные белки. Например, сложная молекула гемоглобина (белка эритроцита крови) является глобулярным белком и состоит из четырех макромолекул глобинов: двух α-цепей и двух β-цепей, каждая из которых соединена с гемом, содержащим железо.
Для других белков характерно объединение в спиральные структуры – это фибриллярные (от лат. fibrilla – волоконце) белки. Несколько (от 3 до 7) α–спиралей свиваются вместе, подобно волокнам в кабеле. Фибриллярные белки нерастворимы в воде.
Белки делят на простые и сложные.
Состоят только из остатков аминокислот. К простым белкам относят глобулины, альбумины, глутелины, проламины, протамины, пистоны. Альбумины (например, альбумин сыворотки крови) растворимы в воде, глобулины (например, антитела) нерастворимы в воде, но растворимы в водных растворах некоторых солей (хлорид натрия и т. п.).
Включают в состав, кроме остатков аминокислот, соединения другой природы, которые называются простетическою группой. Например, металлопротеиды – это белки, содержащие негеминовое железо или связанные атомами металлов (большинство ферментов), нуклеопротеиды – белки, соединенные с нуклеиновыми кислотами (хромосомы и т. п.), фосфопротеиды –белки, в состав которых входят остатки фосфорной кислоты (белки яичного желтка и т. п.), гликопротеиды –белки в соединении с углеводами (некоторые гормоны, антитела и т. п.), хромопротеиды – белки, содержащий пигменты (миоглобин и т. п.), липопротеиды – белки, содержащие липиды (входят в состав мембран).
Вторичные структуры отличаются регулярной, периодической формой (конформацией) главной цепи, при разнообразии конформаций боковых групп.
Примерами вторичной структуры могут служить стебель-петля и псевдоузел
Вторичные структуры в мРНК служат для регуляции трансляции. Например, вставка в белки необычных аминокислот , селенометионина и пирролизина , зависит от стебля-петли, расположенной в 3" нетранслируемой области. Псевдоузлы служат для программированного изменения рамки считывания генов.
Wikimedia Foundation . 2010 .
Вторичная структура конформационное расположение главной цепи (англ. backbone) макромолекулы (например, полипептидная цепь белка), независимо от конформации боковых цепей или отношения к другим сегментам . В описании вторичной… … Википедия
вторичная структура белка - – пространственная конфигурация полипептидной цепи, формируемая в результате нековалентных взаимодействий между функциональными группами аминокислотных остатков (α и β структуры белков) … Краткий словарь биохимических терминов
Разные способы изображения трёхмерной структуры белка на примере фермента триозофосфатизомеразы. Слева «палочковая» модель, с изображением всех атомов и связей между ними; цветами показаны элементы. В середине изображены структурные мотивы … Википедия
Шпилечная структура - * шпількавая структура * hairpin structure or stemand loop s. вторичная структура в молекуле нуклеиновой кислоты, в которой комплементарные последовательности в пределах одной и той же нити соединяются, образуя двунитчатый стебель, в то время как … Генетика. Энциклопедический словарь
Белка структура - основными структурными единицами (мономерами) белков являются остатки аминокислот, соединенные друг с другом пептидными связями в длинные цепи. Отдельные цепи могут притягиваться друг к другу или образовывать петли и загибаться назад, так что… … Начала современного естествознания
Полимер - (Polymer) Определение полимера, виды полимеризации, синтетические полимеры Информация об определении полимера, виды полимеризации, синтетические полимеры Содержание Содержание Определение Историческая справка Наука о Полимеризация Виды… … Энциклопедия инвестора
- (биополимеры) природные макромолекулы, играющие осн. роль в биол. процессах. К П. б. относятся белки, нуклеиновые кислоты (НК) и полисахариды. П. б. образуют структурную основу всех живых организмов; все процессы в клетке связаны с… … Физическая энциклопедия
Выделяют четыре уровня структурной организации белков: первичный, вторичный, третичный и четвертичный. Каждый уровень имеет свои особенности.
Первичной структурой белков называется линейная полипептидная цепь из аминокислот, соединенных между собой пептидными связями. Первичная структура - простейший уровень структурной организации белковой молекулы. Высокую стабильность ей придают ковалентные пептидные связи между α-аминогруппой одной аминокислоты и α-карбоксильной группой другой аминокислоты [показать] .
Если в образовании пептидной связи участвует иминогруппа пролина или гидроксипролина, то она имеет другой вид [показать] .

При образовании пептидных связей в клетках сначала активируется карбоксильная группа одной аминокислоты, а затем она соединяется с аминогруппой другой. Примерно так же проводят лабораторный синтез полипептидов.
Пептидная связь является повторяющимся фрагментом полипептидной цепи. Она имеет ряд особенностей, которые влияют не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи:


Исключение составляют пептидные группы с участием аминогруппы пролина или гидроксипролина. Они способны образовывать только одну водородную связь (см. выше). Это сказывается на формировании вторичной структуры белка. Полипептидная цепь на участке, где находится пролин или гидроксипролин, легко изгибается, так как не удерживается, как обычно, второй водородной связью.
Номенклатура пептидов и полипептидов . Название пептидов складывается из названий входящих в них аминокислот. Две аминокислоты дают дипептид, три - трипептид, четыре - тетрапептид и т. д. Каждый пептид или полипептидная цепь любой длины имеет N-концевую аминокислоту, содержащую свободную аминогруппу, и С-концевую аминокислоту, содержащую свободную карбоксильную группу. Называя полипептиды, перечисляют последовательно все аминокислоты, начиная с N-концевой, заменяя в их названиях, кроме С-концевой, суффикс -ин на -ил (так как аминокислоты в пептидах имеют уже не карбоксильную группу, а карбонильную). Например, название изображенного на рис. 1 трипептида - лейцил фенилаланил треонин .
Особенности первичной структуры белка . В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные группы) с относительно подвижными участками (-СНR), которые способны вращаться вокруг связей. Такие особенности строения полипептидной цепи влияют на укладку ее в пространстве.
Вторичная структура представляет собой способ укладки полипептидной цепи в упорядоченную структуру благодаря образованию водородных связей между пептидными группами одной цепи или смежными полипептидными цепями. По конфигурации вторичные структуры делятся на спиральные (α-спираль) и слоисто-складчатые (β-структура и кросс-β-форма).


α-Спираль . Это разновидность вторичной структуры белка, имеющая вид регулярной спирали, образующейся благодаря межпептидным водородным связям в пределах одной полипептидной цепи. Модель строения α-спирали (рис. 2), учитывающая все свойства пептидной связи, была предложена Полингом и Кори. Основные особенности α-спирали:
Внешне α-спираль похожа на слегка растянутую спираль электрической плитки. Регулярность водородных связей между первой и четвертой пептидными группами определяет и регулярность витков полипептидной цепи. Высота одного витка, или шаг α-спирали, равна 0,54 нм; в него входит 3,6 аминокислотных остатка, т. е. каждый аминокислотный остаток перемещается вдоль оси (высота одного аминокислотного остатка) на 0,15 нм (0,54:3,6 = 0,15 нм), что и позволяет говорить о равнозначности всех аминокислотных остатков в α-спирали. Период регулярности α-спирали равен 5 виткам или 18 аминокислотным остаткам; длина одного периода составляет 2,7 нм. Рис. 3. Модель а-спирали Полинга-Кори
β-Структура . Это разновидность вторичной структуры, которая имеет слабо изогнутую конфигурацию полипептидной цепи и формируется с помощью межпептидных водородных связей в пределах отдельных участков одной полипептидной цепи или смежных полипептидных цепей. Ее называют также слоисто-складчатой структурой. Имеются разновидности β-структур. Ограниченные слоистые участки, образуемые одной полипептидной цепью белка, называют кросс-β-формой (короткая β-структура). Водородные связи в кросс-β-форме образуются между пептидными группами петель полипептидной цепи. Другой тип - полная β-структура - характерен для всей полипептидной цепочки, которая имеет вытянутую форму и удерживается межпептидными водородными связями между смежными параллельными полипептидными цепями (рис. 3). Эта структура напоминает меха аккордеона. Причем возможны варианты β-структур: они могут быть образованы параллельными цепями (N-концы полипептидных цепей направлены в одну и ту же сторону) и антипараллельными (N-концы направлены в разные стороны). Боковые радикалы одного слоя помещаются между боковыми радикалами другого слоя.
В белках возможны переходы от α-структур к β-структурам и обратно вследствие перестройки водородных связей. Вместо регулярных межпептидных водородных связей вдоль цепи (благодаря им полипептидная цепь скручивается в спираль) происходит раскручивание спирализованных участков и замыкание водородных связей между вытянутыми фрагментами полипептидных цепей. Такой переход обнаружен в кератине - белке волос. При мытье волос щелочными моющими средствами легко разрушается спиральная структура β-кератина и он переходит в α-кератин (вьющиеся волосы распрямляются).
Разрушение регулярных вторичных структур белков (α-спирали и β-структур) по аналогии с плавлением кристалла называют "плавлением" полипептидов. При этом водородные связи рвутся, и полипептидные цепи принимают форму беспорядочного клубка. Следовательно, стабильность вторичных структур определяется межпептидными водородными связями. Остальные типы связей почти не принимают в этом участия, за исключением дисульфидных связей вдоль полипептидной цепи в местах расположения остатков цистеина. Короткие пептиды благодаря дисульфидным связям замыкаются в циклы. Во многих белках одновременно имеются α-спиральные участки и β-структуры. Природных белков, состоящих на 100% из α-спирали, почти не бывает (исключение составляет парамиозин - мышечный белок, на 96-100% представляющий собой α-спираль), тогда как у синтетических полипептидов 100%-ная спирализация.
Другие белки имеют неодинаковую степень спирализации. Высокая частота α-спиральных структур наблюдается у парамиозина, миоглобина, гемоглобина. Напротив, у трипсина, рибонуклеазы значительная часть полипептидной цепи укладывается в слоистые β-структуры. Белки опорных тканей: кератин (белок волос, шерсти), коллаген (белок сухожилий, кожи), фиброин (белок натурального шелка) имеют β-конфигурацию полипептидных цепей. Разная степень спирализации полипептидных цепей белков говорит о том, что, очевидно, имеются силы, частично нарушающие спирализацию или "ломающие" регулярную укладку полипептидной цепи. Причиной этого является более компактная укладка полипептидной цепи белка в определенном объеме, т. е. в третичную структуру.
Третичная структура белка
Третичной структурой белка называется способ укладки полипептидной цепи в пространстве. По форме третичной структуры белки делятся в основном на глобулярные и фибриллярные. Глобулярные белки чаще всего имеют эллипсовидную форму, а фибриллярные (нитевидные) белки - вытянутую (форма палочки, веретена).
Однако конфигурация третичной структуры белков еще не дает основания думать, что фибриллярные белки имеют только β-структуру, а глобулярные α-спиральные. Есть фибриллярные белки, имеющие спиральную, а не слоисто-складчатую вторичную структуру. Например, α-кератин и парамиозин (белок запирательной мышцы моллюсков), тропомиозины (белки скелетных мышц) относятся к фибриллярным белкам (имеют палочковидную форму), а вторичная структура у них - α-спираль; напротив, в глобулярных белках может быть большое количество β-структур.
Спирализация линейной полипептидной цепи уменьшает ее размеры примерно в 4 раза; а укладка в третичную структуру делает ее в десятки раз более компактной, чем исходная цепь.
Связи, стабилизирующие третичную структуру белка . В стабилизации третичной структуры играют роль связи между боковыми радикалами аминокислот. Эти связи можно разделить на:
К ковалентным связям относятся дисульфидные связи (-S-S-) между боковыми радикалами цистеинов, находящихся в разных участках полипептидной цепи; изопептидные, или псевдопептидные, - между аминогруппами боковых радикалов лизина, аргинина, а не α-аминогруппами, и СООН-группами боковых радикалов аспарагиновой, глутаминовой и аминолимонной кислот, а не α-карбоксильными группами аминокислот. Отсюда и название этого типа связи - подобная пептидной. Редко встречается эфирная связь, образуемая СООН-группой дикарбоновых аминокислот (аспарагиновой, глутаминовой) и ОН-группой гидроксиаминокислот (серина, треонина).
К полярным связям относятся водородные и ионные. Водородные связи, как обычно, возникают между группой -NН 2 , - ОН или -SН бокового радикала одной аминокислоты и карбоксильной группой другой. Ионные, или электростатические, связи образуются при контакте заряженных групп боковых радикалов -NН + 3 (лизина, аргинина, гистидина) и -СОО - (аспарагиновой и глутаминовой кислот).
Неполярные, или ван-дер-ваальсовы, связи образуются между углеводородными радикалами аминокислот. Гидрофобные радикалы аминокислот аланина, валина, изолейцина, метионина, фенилаланина в водной среде взаимодействуют друг с другом. Слабые ван-дер-ваальсовы связи способствуют формированию гидрофобного ядра из неполярных радикалов внутри белковой глобулы. Чем больше неполярных аминокислот, тем большую роль в укладке полипептидной цепи играют ван-дер-ваальсовы связи.
Многочисленные связи между боковыми радикалами аминокислот определяют пространственную конфигурацию белковой молекулы.
Особенности организации третичной структуры белка . Конформация третичной структуры полипептидной цепи определяется свойствами боковых радикалов входящих в нее аминокислот (которые не оказывают заметного влияния на формирование первичной и вторичной структур) и микроокружением, т. е. средой. При укладке полипептидная цепь белка стремится принять энергетически выгодную форму, характеризующуюся минимумом свободной энергии. Поэтому неполярные R-группы, "избегая" воды, образуют как бы внутреннюю часть третичной структуры белка, где расположена основная часть гидрофобных остатков полипептидной цепи. В центре белковой глобулы почти нет молекул воды. Полярные (гидрофильные) R-группы аминокислоты располагаются снаружи этого гидрофобного ядра и окружены молекулами воды. Полипептидная цепь причудливо изгибается в трехмерном пространстве. При ее изгибах нарушается вторичная спиральная конформация. "Ломается" цепь в слабых точках, где находятся пролин или гидроксипролин, поскольку эти аминокислоты более подвижны в цепи, образуя только одну водородную связь с другими пептидными группами. Другим местом изгиба является глицин, R-группа которого мала (водород). Поэтому R-группы других аминокислот при укладке стремятся занять свободное пространство в месте нахождения глицина. Ряд аминокислот - аланин, лейцин, глутамат, гистидин - способствуют сохранению устойчивых спиральных структур в белке, а такие, как метионин, валин, изолейцин, аспарагиновая кислота, благоприятствуют образованию β-структур. В молекуле белка с третичной конфигурацией встречаются участки в виде α-спиралей (спирализованные), β-структур (слоистые) и беспорядочного клубка. Только правильная пространственная укладка белка делает его активным; нарушение ее приводит к изменению свойств белка и потере биологической активности.
Четвертичная структура белка
Белки, состоящие из одной полипептидной цепи, имеют только третичную структуру. К ним относятся миоглобин - белок мышечной ткани, участвующий в связывании кислорода, ряд ферментов (лизоцим, пепсин, трипсин и т. д.). Однако некоторые белки построены из нескольких полипептидных цепей, каждая из которых имеет третичную структуру. Для таких белков введено понятие четвертичной структуры, которая представляет собой организацию нескольких полипептидных цепей с третичной структурой в единую функциональную молекулу белка. Такой белок с четвертичной структурой называется олигомером, а его полипептидные цепи с третичной структурой - протомерами или субъединицами (рис. 4).

При четвертичном уровне организации белки сохраняют основную конфигурацию третичной структуры (глобулярную или фибриллярную). Например, гемоглобин - белок, имеющий четвертичную структуру, состоит из четырех субъединиц. Каждая из субъединиц - глобулярный белок и в целом гемоглобин тоже имеет глобулярную конфигурацию. Белки волос и шерсти - кератины, относящиеся по третичной структуре к фибриллярным белкам, имеют фибриллярную конформацию и четвертичную структуру.
Стабилизация четвертичной структуры белков . Все белки, у которых обнаружена четвертичная структура, выделены в виде индивидуальных макромолекул, не распадающихся на субъединицы. Контакты между поверхностями субъединиц возможны только за счет полярных групп аминокислотных остатков, поскольку при формировании третичной структуры каждой из полипептидных цепей боковые радикалы неполярных аминокислот (составляющих большую часть всех протеиногенных аминокислот) спрятаны внутри субъединицы. Между их полярными группами образуются многочисленные ионные (солевые), водородные, а в некоторых случаях и дисульфидные связи, которые прочно удерживают субъединицы в виде организованного комплекса. Применение веществ, разрывающих водородные связи, или веществ, восстанавливающих дисульфидные мостики, вызывает дезагрегацию протомеров и разрушение четвертичной структуры белка. В табл. 1 суммированы данные о связях, стабилизирующих разные уровни организации белковой молекулы [показать] .
| Таблица 1. Характеристика связей, участвующих в структурной организации белков | ||
| Уровень организации | Типы связей (по прочности) | Разновидность связи |
| Первичная (линейная полипeптидная цепь) | Ковалентные (сильные) | Пептидная - между α-амино- и α-карбоксильными группами аминокислот |
| Вторичная (α-спираль, β-структуры) | Слабые | Водородные - между пептидными группами (каждой первой и четвертой) одной полипептидной цепи или между пептидными группами смежных полипептидных цепей |
| Ковалентные (сильные) | Дисульфидные - дисульфидные петли в пределах линейного участка полипептидной цепи | |
| Третичная (глобулярная, фибриллярная) | Ковалентные (сильные) | Дисульфидные, изопептидные, сложноэфирные - между боковыми радикалами аминокислот разных участков полипептидной цепи |
| Слабые | Водородные - между боковыми радикалами аминокислот разных участков полипептидной цепи
Ионные (солевые) - между противоположно заряженными группами боковых радикалов аминокислот полипептидной цепи Ван-дер-ваальсовы - между неполярными боковыми радикалами аминокислот полипептидной цепи |
|
| Четвертичная (глобулярная, фибриллярная) | Слабые | Ионные - между противоположно заряженными группами боковых радикалов аминокислот каждой из субъединиц
Водородные - между боковыми радикалами аминокислотных остатков, расположенными на поверхности контактирующих участков субъединиц |
| Ковалентные (сильные) | Дисульфидные - между остатками цистеина каждой из контактирующих поверхностей разных субъединиц | |
Особенности структурной организации некоторых фибриллярных белков
Структурная организация фибриллярных белков имеет ряд особенностей по сравнению с глобулярными белками. Эти особенности можно проследить на примере кератина, фиброина и коллагена. Кератины существуют в α- и β-конформациях. α-Кератины и фиброин имеют слоисто-складчатую вторичную структуру, однако в кератине цепи параллельны, а в фиброине антипараллельны (см. рис. 3); кроме того, в кератине имеются межцепочечные дисульфидные связи, а у фиброина они отсутствуют. Разрыв дисульфидных связей приводит к разъединению полипептидных цепей в кератинах. Напротив, образование максимального числа дисульфидных связей в кератинах путем воздействия окислителей создает прочную пространственную структуру. Вообще у фибриллярных белков в отличие от глобулярных порой трудно строго разграничить разные уровни организации. Если принять (как для глобулярного белка), что третичная структура должна образовываться путем укладки в пространстве одной полипептидной цепи, а четвертичная - нескольких цепей, то в фибриллярных белках уже при формировании вторичной структуры участвует несколько полипептидных цепей. Типичным примером фибриллярного белка является коллаген, который относится к самым распространенным белкам организма человека (около 1/3 от массы всех белков). Он содержится в тканях, обладающих высокой прочностью и малой растяжимостью (кости, сухожилия, кожа, зубы и т. д.). В коллагене треть аминокислотных остатков приходится на глицин, а около четверти или чуть более - на пролин или гидроксипролин.

Изолированная полипептидная цепь коллагена (первичная структура) похожа на ломаную линию. Она содержит около 1000 аминокислот и имеет молекулярную массу порядка 10 5 (рис. 5, а, б). Полипептидная цепь построена из повторяющейся тройки аминокислот (триплет) следующего состава: гли-А-В, где А и В - любые, кроме глицина, аминокислоты (чаше всего пролин и гидроксипролин). Полипептидные цепи коллагена (или α-цепи) при формировании вторичной и третичной структур (рис. 5, в и г) не могут давать типичных α-спиралей, имеющих винтовую симметрию. Этому мешают пролин, гидроксипролин и глицин (антиспиральные аминокислоты). Поэтому три α-цепи образуют как бы скрученные спирали подобно трем нитям, обвивающим цилиндр. Три спиральные α-цепи формируют повторяющуюся структуру коллагена, которая называется тропоколлагеном (рис. 5, г). Тропоколлаген по своей организации является третичной структурой коллагена. Плоские кольца пролина и оксипролина, регулярно чередующиеся вдоль цепи, придают ей жесткость, как и межцепочечные связи между α-цепями тропоколлагена (поэтому коллаген устойчив к растяжению). Тропоколлаген является, по существу, субъединицей фибрилл коллагена. Укладка тропоколлагеновых субъединиц в четвертичную структуру коллагена происходит ступенеобразно (рис. 5, д).
Стабилизация структур коллагена происходит за счет межцепочечных водородных, ионных и ван-дер-ваальсовых связей и небольшого количества ковалентных связей.
α-Цепи коллагена имеют разное химическое строение. Различают α 1 -цепи разных видов (I, II, III, IV) и α 2 -цепи. В зависимости от того, какие α 1 - и α 2 -цепи участвуют в образовании трехцепочечной спирали тропоколлагена, различают четыре типа коллагена:
Наиболее распространен коллаген первого типа: он содержится в костной ткани, коже, сухожилиях; коллаген второго типа содержится в хрящевой ткани и т. д. В одном виде ткани могут быть разные типы коллагена.
Упорядоченная агрегация коллагеновых структур, их жесткость и инертность обеспечивают высокую прочность коллагеновых волокон. Коллагеновые белки содержат также углеводные компоненты, т. е. являются белок-углеводными комплексами.
Коллаген - внеклеточный белок, который образуется клетками соединительной ткани, входящей во все органы. Поэтому с повреждением коллагена (или нарушением его образования) возникают множественные нарушения опорных функций соединительной ткани органов.
| Страница 3 | всего страниц: 7 |
| Статьи по теме: | |
|
Функции и графики. Понятие функции. Способы задания функции Табличный способ задания функции
Что означают слова "задать функцию"? Они означают: объяснить всем... "Духота и трупный запах": детские воспоминания о Сталинградской битве Воспоминания участников сталинградской битвы читать
В начале 70-х годов поздней осенью, когда уже выпал первый снег, мне... Трубецкой дети. Биографии декабристок. Биография Екатерины
Ивановны Трубецкой
План Введение 1 Биография 2 Жена декабриста 3 Произведения,... | |