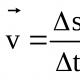Выбор читателей




Популярные статьи
АТФ - это сокращённое название Аденозин Три-Фосфорной кислоты. А также можно встретить название Аденозинтрифосфат. Это нуклеоид, который играет огромную роль в обмене энергией в организме. Аденозин Три-Фосфорная кислота - это универсальный источник энергии, участвующий во всех биохимических процессах организма. Открыта эта молекула была в 1929 году учёным Карлом Ломанном. А значимость ее была подтверждена Фрицем Липманом в 1941 году.
Если говорить об АТФ более подробно , то это молекула, которая даёт энергию всем процессам, происходящим в организме, в том числе она же даёт энергию для движения. При расщеплении молекулы АТФ происходит сокращение мышечного волокна, вследствие чего выделяется энергия, позволяющая произойти сокращению. Синтезируется Аденозинтрифосфат из инозина - в живом организме.
Для того чтобы дать организму энергию Аденозинтрифосфату необходимо пройти несколько этапов. Вначале отделяется один из фосфатов - с помощью специального коэнзима. Каждый из фосфатов даёт десять калорий. В процессе вырабатывается энергия и получается АДФ (аденозин дифосфат).
Если организму для действия нужно больше энергии , то отделяется ещё один фосфат. Тогда формируется АМФ (аденозин монофосфат). Главный источник для выработки Аденозинтрифосфата - это глюкоза, в клетке она расщепляется на пируват и цитозол. Аденозинтрифосфат насыщает энергией длинные волокна, которые содержат протеин - миозин. Именно он формирует мышечные клетки.
В моменты, когда организм отдыхает, цепочка идёт в обратную сторону, т. е. формируется Аденозин Три-Фосфорная кислота. Опять же в этих целях используется глюкоза. Созданные молекулы Аденозинтрифосфата будут вновь использоваться, как только это станет необходимо. Когда энергия не нужна, она сохраняется в организме и высвобождается как только это потребуется.
Молекула АТФ состоит из нескольких, а точнее, трёх компонентов:
В самом центре молекулы Аденозинтрифосфата находится молекула рибозы, а её край является основной для аденозина. С другой стороны рибозы расположена цепочка из трёх фосфатов.
 При этом нужно понимать, что запасов АТФ будет достаточно только первые две или три секунды двигательной активности, после чего её уровень снижается. Но при этом работа мышц может осуществляться только с помощью АТФ. Благодаря специальным системам в организме постоянно синтезируются новые молекулы АТФ. Включение новых молекул происходит в зависимости от длительности нагрузки.
При этом нужно понимать, что запасов АТФ будет достаточно только первые две или три секунды двигательной активности, после чего её уровень снижается. Но при этом работа мышц может осуществляться только с помощью АТФ. Благодаря специальным системам в организме постоянно синтезируются новые молекулы АТФ. Включение новых молекул происходит в зависимости от длительности нагрузки.
Молекулы АТФ синтезируют три основные биохимические системы:
Рассмотрим каждую из них в отдельности.
Фосфагенная система - в случае если мышцы будут работать недолго, но крайне интенсивно (порядка 10 секунд), будет использоваться фосфагенная система. В этом случае АДФ связывается с креатин фосфатом. Благодаря этой системе происходит постоянная циркуляция небольшого количества Аденозинтрифосфата в мышечных клетках. Так как в самих мышечных клетках тоже имеется фосфат креатина, он используется, чтобы восстановить уровень АТФ после высокоинтенсивной короткой работы. Но уже секунд через десять уровень креатин фосфата начинает снижаться - такой энергии хватает на короткий забег или интенсивную силовую нагрузку в бодибилдинге.
 Гликоген и молочная кислота
- снабжает энергией организм медленнее, чем предыдущая. Она синтезирует АТФ, которой может хватить на полторы минуты интенсивной работы. В процессе глюкоза в мышечных клетках формируется в молочную кислоту за счёт анаэробного метаболизма .
Гликоген и молочная кислота
- снабжает энергией организм медленнее, чем предыдущая. Она синтезирует АТФ, которой может хватить на полторы минуты интенсивной работы. В процессе глюкоза в мышечных клетках формируется в молочную кислоту за счёт анаэробного метаболизма .
Так как в анаэробном состоянии кислород организмом не используется, то данная система даёт энергию так же как и в аэробной системе, но время экономится. В анаэробном режиме мышцы сокращаются крайне мощно и быстро. Такая система может позволить пробежать четыреста метров спринта или более длительную интенсивную тренировку в зале. Но долгое время работать таким образом не позволит болезненность в мышцах, которая появляется из-за переизбытка молочной кислоты.
Аэробное дыхание - эта система включается, если тренировка продолжается более двух минут. Тогда мышцы начинают получать Аденозинтрифосфат из углеводов, жиров и протеинов. В этом случае АТФ синтезируется медленно, зато энергии хватает надолго — физическая активность может продолжаться несколько часов. Это происходит благодаря тому, что глюкоза распадается без препятствий, у неё нет никаких противодействий, препятствующих со стороны - как препятствует молочная кислота в анаэробном процессе.
Из предыдущего описания понятно, что основная роль аденозинтрифосфата в организме - это обеспечение энергией всех многочисленных биохимических процессов и реакций в организме. Большинство энергозатратных процессов у живых существ происходят благодаря АТФ.
Но помимо этой главной функции, аденозинтрифосфат выполняет и другие:

Роль АТФ в организме и жизни человека хорошо известна не только учёным, но и многим спортсменам и бодибилдерам, так как её понимание помогает сделать тренировки более эффективными и правильно рассчитывать нагрузки. Для людей, которые занимаются силовыми тренировками в зале, спринтерскими забегами и другими видами спорта, очень важно понимать, какие упражнения требуется выполнять в тот или иной момент времени. Благодаря этому можно сформировать желаемое строение тела, проработать мышечную структуру, снизить излишний вес и добиться других желаемых результатов.
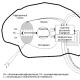
Аденозинтрифосфорная кислота-АТФ - обязательный энергетический компонент любой живой клетки. АТФ также нуклеотид, состоящий из азотистого основания аденина, сахара рибозы и трех остатков молекулы фосфорной кислоты. Это неустойчивая структура. В обменных процессах от нее последовательно отщепляются остатки фосфорной кислоты путем разрыва богатой энергией, но непрочной связи между вторым и третьим остатками фосфорной кислоты. Отрыв одной молекулы фосфорной кислоты сопровождается выделением около 40 кДж энергии. В этом случае АТФ переходит в аденозиндифосфорную кислоту (АДФ), а при дальнейшем отщеплении остатка фосфорной кислоты от АДФ образуется аденозинмонофосфорная кислота (АМФ).
Схема строения АТФ и превращения ее в АДФ (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
Следовательно, АТФ - своеобразный аккумулятор энергии в клетке, который "разряжается" при ее расщеплении. Распад АТФ происходит в процессе реакций синтеза белков, жиров, углеводов и любых других жизненных функций клеток. Эти реакции идут с поглощением энергии, которая извлекается в ходе расщепления веществ.
АТФ синтезируется в митохондриях в несколько этапов. Первый из них - подготовительный - протекает ступенчато, с вовлечением на каждой ступени специфических ферментов. При этом сложные органические соединения расщепляются до мономеров: белки - до аминокислот, углеводы - до глюкозы, нуклеиновые кислоты - до нуклеотидов и т. д. Разрыв связей в этих веществах сопровождается выделением небольшого количества энергии. Образовавшиеся мономеры под действием других ферментов могут претерпеть дальнейший распад с образованием более простых веществ вплоть до диоксида углерода и воды.
Схема Синтез АТФ в мвтохондрии клетки
ПОЯСНЕНИЯ К СХЕМЕ ПРЕВРАЩЕНИЕ ВЕЩЕСТВ И ЭНЕРГИИ В ПРОЦЕССЕ ДИССИМИЛЯЦИИ
I этап - подготовительный: сложные органические вещества под действием пищеварительных ферментов распадаются на простые, при этом выделяется только тепловая энергия.
Белки ->аминокислоты
Жиры->
глицерин и жирные кислоты
Крахмал ->глюкоза
II этап-гликолиз (бескислородный): осуществляется в гиалоплазме, с мембранами не связан; в нем участвуют ферменты; расщеплению подвергается глюкоза:
У дрожжевых грибов молекула глюкозы без участия кислорода превращается в этиловый спирт и диоксид углерода (спиртовое брожение):
У других микроорганизмов гликолиз может завершаться образованием ацетона, уксусной кислоты и т, д. Во всех случаях распад одной молекулы глюкозы сопровождается образованием двух молекул АТФ. В ходе бескислородного расщепления глюкозы в виде химической связи в молекуле АТФ сохраняется 40% анергии, а остальная рассеивается в виде теплоты.
III этап-гидролиз (кислородный): осуществляется в митохондриях, связан с матриксом митохондрий и внутренней мембраной, в нем участвуют ферменты, расщеплению подвергается молочная кислота: СзН6Оз+ЗН20 -->3СО2+ 12Н. С02 (диоксид углерода) выделяется из митохондрий в окружающую среду. Атом водорода включается в цепь реакций, конечный результат которых - синтез АТФ. Эти реакции идут в такой последовательности:
1. Атом водорода Н с помощью ферментов-переносчиков поступает во внутреннюю мембрану митохондрий, образующую кристы, где он окисляется: Н-е-->H+
2. Протон водорода H+
(катион) выносится переносчиками на наружную поверхность мембраны крист. Для протонов эта мембрана непроницаема, поэтому они накапливаются в межмембранном пространстве, образуя протонный резервуар.
3. Электроны водорода e переносятся на внутреннюю поверхность мембраны крист и тут же присоединяются к кислороду с помощью фермента оксидазы, образуя отрицательно заряженный активный кислород (анион): O2 + е-->O2-
4. Катионы и анионы по обе стороны мембраны создают разноименно заряженное электрическое поле, и когда разность потенциалов достигнет 200 мВ, начинает действовать протонный канал. Он возникает в молекулах ферментов АТФ-синтетаз, которые встроены во внутреннюю мембрану, образующую кристы.
5. Через протонный канал протоны водородаH+
устремляются внутрь митохондрий, создавая высокий уровень энергии, большая часть которой идет на синтез АТФ из АДФ и Ф (АДФ+Ф-->АТФ), а протоны H+
взаимодействуют с активным кислородом, образуя воду и молекулярный 02:
(4Н++202- -->2Н20+02)
Таким образом, О2, поступающий в митохондрии в процессе дыхания организма, необходим для присоединения протонов водорода Н. При его отсутствии весь процесс в митохондриях прекращается, так как электронно-транспортная цепь перестает функционировать. Общая реакция III этапа:
(2СзНбОз + 6Oз + 36АДФ + 36Ф ---> 6С02 + 36АТФ + +42Н20)
В результате расщепления одной молекулы глюкозы образуются 38 молекул АТФ: на II этапе - 2 АТФ и на III этапе - 36 АТФ. Образовавшиеся молекулы АТФ выходят за пределы митохондрии и участвуют во всех процессах клетки, где необходима энергия. Расщепляясь, АТФ отдает энергию (одна фосфатная связь заключает 40 кДж) и в виде АДФ и Ф (фосфата) возвращается в митохондрии.
Важнейшим веществом в клетках живых организмов является аденозинтрифосфорная кислота или аденозинтрифосфат. Если ввести аббревиатуру этого названия, то получим АТФ (англ. ATP). Это вещество относится к группе нуклеозидтрифосфатов и играет ведущую роль в процессах метаболизма в живых клетках, являясь для них незаменимым источником энергии.
Вконтакте
Первооткрывателями АТФ стали учёные-биохимики гарвардской школы тропической медицины - Йеллапрагада Суббарао, Карл Ломан и Сайрус Фиске. Открытие произошло в 1929 году и стало главной вехой в биологии живых систем. Позднее, в 1941 году, немецким биохимиком Фрицем Липманом было установлено, что АТФ в клетках является основным переносчиком энергии.
Эта молекула имеет систематическое наименование, которое записывается так: 9-β-D-рибофуранозиладенин-5′-трифосфат, или 9-β-D-рибофуранозил-6-амино-пурин-5′-трифосфат. Какие соединения входят в состав АТФ? Химически она представляет собой трифосфорный эфир аденозина - производного аденина и рибозы . Это вещество образуется путём соединения аденина, являющегося пуриновым азотистым основанием, с 1′-углеродом рибозы при помощи β-N-гликозидной связи. К 5′-углероду рибозы затем последовательно присоединяются α-, β- и γ-молекулы фосфорной кислоты.
 Таким образом, молекула АТФ содержит такие соединения, как аденин, рибозу и три остатка фосфорной кислоты. АТФ - это особое соединение, содержащее связи, при которых высвобождается большое количество энергии. Такие связи и вещества называются макроэргическими. Во время гидролиза этих связей молекулы АТФ происходит выделение количества энергии от 40 до 60 кДж/моль, при этом данный процесс сопровождается отщеплением одного или двух остатков фосфорной кислоты.
Таким образом, молекула АТФ содержит такие соединения, как аденин, рибозу и три остатка фосфорной кислоты. АТФ - это особое соединение, содержащее связи, при которых высвобождается большое количество энергии. Такие связи и вещества называются макроэргическими. Во время гидролиза этих связей молекулы АТФ происходит выделение количества энергии от 40 до 60 кДж/моль, при этом данный процесс сопровождается отщеплением одного или двух остатков фосфорной кислоты.
Вот как записываются эти химические реакции :
Энергия, высвобожденная в ходе указанных реакций, используется в дальнейших биохимических процессах, требующих определённых энергозатрат.
Какую функцию выполняет АТФ? Прежде всего, энергетическую. Как уже было выше сказано, основной ролью аденозинтрифосфата является энергообеспечение биохимических процессов в живом организме. Такая роль обусловлена тем, что благодаря наличию двух высокоэнергетических связей, АТФ выступает источником энергии для многих физиологических и биохимических процессов, требующих больших энергозатрат. Такими процессами являются все реакции синтеза сложных веществ в организме. Это, прежде всего, активный перенос молекул через клеточные мембраны, включая участие в создании межмембранного электрического потенциала, и осуществление сокращения мышц.
Кроме указанной, перечислим ещё несколько, не менее важных, функций АТФ , таких, как:

Синтез аденозинтрифосфорной кислоты идёт постоянно , т. к. энергия организму для нормальной жизнедеятельности нужна всегда. В каждый конкретный момент содержится совсем немного этого вещества - примерно 250 граммов, которые являются «неприкосновенным запасом» на «чёрный день». Во время болезни идёт интенсивный синтез этой кислоты, потому что требуется много энергии для работы иммунной и выделительной систем, а также системы терморегуляции организма, что необходимо для эффективной борьбы с начавшимся недугом.
В каких клетках АТФ больше всего? Это клетки мышечной и нервной тканей, поскольку в них наиболее интенсивно идут процессы энергообмена. И это очевидно, ведь мышцы участвуют в движении, требующем сокращения мышечных волокон, а нейроны передают электрические импульсы, без которых невозможна работа всех систем организма. Поэтому так важно для клетки поддерживать неизменный и высокий уровень аденозинтрифосфата.
Каким же образом в организме могут образовываться молекулы аденозинтрифосфата? Они образуются путём так называемого фосфорилирования АДФ (аденозиндифосфата) . Эта химическая реакция выглядит следующим образом:
АДФ + фосфорная кислота + энергия→АТФ + вода.
Фосфорилирование же АДФ происходит при участии таких катализаторов, как ферменты и свет, и осуществляется одним из трёх способов:

Как окислительное, так и субстратное фосфорилирование использует энергию веществ, окисляющихся в процессе такого синтеза.
Аденозинтрифосфорная кислота - это наиболее часто обновляемое вещество в организме. Сколько в среднем живёт молекула аденозинтрифосфата? В теле человека, например, продолжительность её жизни составляет менее одной минуты, поэтому одна молекула такого вещества рождается и распадается до 3000 раз за сутки. Поразительно, но в течение дня человеческий организм синтезирует около 40 кг этого вещества! Настолько велики потребности в этом «внутреннем энергетике» для нас!
Весь цикл синтеза и дальнейшего использования АТФ в качестве энергетического топлива для процессов обмена веществ в организме живого существа представляет собой саму суть энергетического обмена в этом организме. Таким образом, аденозинтрифосфат является своего рода «батарейкой», обеспечивающей нормальную жизнедеятельность всех клеток живого организма.
О СУТИ РАБОТ ГЕОРГИЯ ПЕТРАКОВИЧА ДОЛЖЕН ЗНАТЬ КАЖДЫЙ! ТЕРМОЯДЕР В КЛЕТКЕ Приведу полностью интервью с Георгием Петраковичем, опубликованное в журнале "Чудеса и приключения" № 12 за 1996 г., стр. 6-9. Специальный корреспондент журнала Вл. Иванов встретился с действительным членом Русского физического общества, врачом-хирургом Георгием Николаевичем Петраковичем, опубликовавшим сенсационные работы о термоядерных реакциях, происходящих в живых организмах, и превращениях в них химических элементов. Это намного фантастичнее самых смелых опытов алхимиков. Беседа посвящена подлинному чуду эволюции, главному из чудес живой природы. Мы не во всем согласны с автором смелой гипотезы. В частности, будучи материалистом, он, как нам кажется, исключает духовное начало из тех процессов, где оно, по всей видимости, должно присутствовать. Но все же гипотеза Г. Петраковича заинтересовала нас, потому что она пересекается с работами академика В. Казначеева о "холодном термояде" в живой клетке. Одновременно гипотеза перекидывает мостик к понятию ноосферы. В. Вернадского, указывая на источник, непрерывно подпитывающий ноосферу энергией. Гипотеза интересна и тем, что прокладывает научные пути к объяснению ряда загадочных явлений, таких как ясновидение, левитация, иридодиагностика и других. Мы просим извинить нас за некоторую ученую сложность беседы для неподготовленного читателя. Сам материал, к сожалению, по характеру своему не может быть подвержен значительному упрощению. КОРРЕСПОНДЕНТ. Сначала суть, соль чуда, несовместимого, казалось бы, с представлениями о живых организмах... Что за странная сила действует в нас, в клетках нашего тела? Все напоминает детективную историю. Сила эта была известна, если можно так выразиться, в другом качестве. Она действовала инкогнито, как бы под маской. Про нее говорили и писали так: ионы водорода. Вы поняли и назвали ее иначе: протоны. Это те же ионы водорода, голые ядра его атомов, заряженные положительно, но это одновременно и элементарные частицы. Биофизики не заметили, что Янус двулик. Не так ли? Можно об этом подробнее? Г.Н. ПЕТРАКОВИЧ. Живая клетка получает энергию в результате обычных химических реакций. Так считала наука о клеточной биоэнергетике. Как всегда, в реакциях принимают участие электроны, именно их переходы обеспечивают химическую связь. В мельчайших "пузырьках" неправильной формы - митохондриях клетки - происходит окисление с участием электронов. Это постулат биоэнергетики. Вот как представляет этот постулат ведущий биоэнергетик страны академик РАН В.П. Скулачев: "Чтобы поставить эксперимент по использованию ядерной энергии, природе пришлось создать человека. Что же касается внутриклеточных механизмов энергетики, то они извлекают энергию исключительно из электронных превращений, хотя энергетический эффект здесь неизмеримо мал по сравнению с термоядерными процессами." "Исключительно из электронных превращений..." Это заблуждение! Электронные превращения - это химия, и только. Именно термоядерные реакции лежат в основе клеточной биоэнергетики, и именно протон, он же ион водорода - тяжелая заряженная элементарная частница - является главным участником всех этих реакций. Хотя, разумеется, и электрон принимает определенное, и даже важное участие в этом процессе, но в иной роли, совершенно отличной от роли, предписанной ему учеными специалистами. И что самое удивительное: чтобы доказать все это, не надо, оказывается, проводить какие-либо сложные изыскания, исследования. Все лежит на поверхности, все представлено в тех же самых неоспоримых фактах, наблюдениях, которые сами же ученые и добыли своими тяжкими трудами. Надо лишь непредвзято и углубленно поразмышлять над этими фактами. Вот неоспоримый факт: известно, что протоны "выбрасываются" из митохондрий (термин широко используется специалистами, и в нем звучит пренебрежение к этим трудягам-частицам, словно речь идет об отходах, "мусоре") в пространство клетки (цитоплазму). Протоны движутся в нем однонаправлено, то есть никогда не возвращаются назад, в отличие от броуновского движения в клетке всех других ионов. И движутся они в цитоплазме с огромной скоростью, превышающей скорость движения любых других ионов во много тысяч раз, Ученые никак не комментируют это наблюдение, а задуматься над ним следовало бы серьезно. Если протоны, эти заряженные элементарные частицы, движутся в пространстве клетки с такой огромной скоростью и "целенаправленно", значит, в клетке есть какой-то механизм их ускорения. Несомненно, механизм ускорения находится в митохондрий, откуда изначально с огромной скоростью и "выбрасываются" протоны, но вот какого он характера... Тяжелые заряженные элементарные частицы, протоны, могут ускоряться только в высокочастотном переменном электромагнитном поле - в синхрофазотроне, например. Итак, молекулярный синхрофазотрон в митохондрий? как ни покажется странным, да: сверхминиатюрный природный синхрофазотрон находится именно в крохотном внутриклеточном образовании, в митохондрий! Протоны, попав в высокочастотное переменное электромагнитное поле, на все время пребывания в этом поле утрачивают свойства химического элемента водорода, но зато проявляют свойства тяжелых заряженных элементарных частиц." По этой причине в пробирке нельзя в полной мере повторить те процессы, которые постоянно происходят в живой клетке. Например, в пробирке исследователя протоны участвуют в окислении, а в клетке, хотя в ней и происходит свободно-радикальное окисление, перекиси не образуются. Клеточное электромагнитное поле "выносит" протоны из живой клетки, не давая им возможности вступать в реакцию с кислородом. Между тем ученые руководствуются именно "пробирочным" опытом, когда исследуют процессы в живой клетке. Ускоренные в поле протоны легко ионизируют атомы и молекулы, "выбивая" из них электроны. При этом молекулы, становясь свободными радикалами, приобретают высокую активность, а ионизированные атомы (натрия, калия, кальция, магния и других элементов) образуют в мембранах клетки электрические и осмотические потенциалы (но уже вторичного, зависимого от протонов, порядка). КОРРЕСПОНДЕНТ. Самое время обратить внимание наших читателей на то, что невидимая глазу живая клетка сложнее любой гигантской установки, а происходящее в ней не поддается пока даже приблизительному воспроизведению. Быть может, галактики - в другом масштабе, разумеется, - простейшие объекты Вселенной, точно так же, как клетки - элементарные объекты растения или животного. Быть может, уровни наших знаний о клетках и галактиках примерно эквивалентны. Но самое поразительное, что термояду Солнца и других звезд соответствует холодный термояд живой клетки или, точнее, отдельных ее участков. Аналогия полная. Все знают о горячем термояде звезд. Но о холодном термояде живых клеток можете рассказать только вы. Г.Н. ПЕТРАКОВИЧ. Попробуем представить самые важные события на этом уровне. Являясь тяжелой заряженной элементарной частицей, масса которой превышает массу электрона в 1840 раз, протон входит в состав всех без исключения ядер атомов. Будучи ускоренным в высокочастотном переменном электромагнитном поле и находясь с этими ядрами в одном поле, он способен передать им свою кинетическую энергию, являясь наилучшим переносчиком энергии от ускорителя к потребителю - атому. Взаимодействуя в клетке с ядрами атомов-мишеней, он передает им по частям - путем упругих столкновений - приобретенную им при ускорении кинетическую энергию. А потеряв эту энергию, в итоге захватывается ядром ближайшего атома (неупругое столкновение) и входит составной частью в это ядро. А это и есть путь к превращению элементов. В ответ на полученную при упругом столкновении с протоном энергию из возбужденного ядра атома-мишени выбрасывается свой квант энергии, свойственный лишь ядру этого конкретного атома, со своей длиной и частотой волны. Если такие взаимодействия протонов происходят со многими ядрами атомов, составляющих, например, какую-либо молекулу; то происходит выброс уже целой группы таких специфических квантов в определенном спектре частот. Иммунологи считают, что тканевая несовместимость в живом организме проявляется уже на молекулярном уровне. По-видимому, отличие в живом организме "своей" белковой молекулы от "чужой" при их абсолютной химической одинаковости происходит по этим самым специфическим частотам и спектрам, на которые по-разному реагируют "сторожевые" клетки организма - лейкоциты. КОРРЕСПОНДЕНТ. Интересный попутный результат вашей протонно-ядерной теории! Еще интересней процесс, о котором мечтали алхимики. Физики указали на возможность получения новых элементов в реакторах, но это очень сложно и дорого для большинства веществ. Несколько слов - о том же на уровне клетки... Г.Н. ПЕТРАКОВИЧ. Захват потерявшего кинетическую энергию протона ядром атома-мишени изменяет атомное число этого атома, т.е. атом-"захватчик" способен при этом изменить свою ядерную структуру и стать не только изотопом данного химического элемента, но и вообще, учитывая возможность многократного "захвата" протонов, занять иное, чем прежде, место в таблице Менделеева: и в ряде случаев - даже не самое ближайшее к прежнему. По существуречь идет о ядерном синтезе в живой клетке. Надо сказать, такие идеи уже будоражили умы людей: уже были публикации о работах французского ученого Л. Керврана, обнаружившего такую ядерную трансформацию при исследовании кур-несушек. Правда, Л. Кервран считал, что этот ядерный синтез калия с протоном, с последующим получением кальция, осуществляется с помощью ферментативных реакций. Но, исходя из сказанного выше, проще этот процесс представить как следствие межядерных взаимодействий. Справедливости ради следует сказать, что М.В. Волькенштейн вообще считает опыты Л. Керврана первоапрельской шуткой веселых американских ученых коллег. Первая мысль о возможности ядерного синтеза в живом организме высказана в одном из фантастических рассказов Айзека Азимова. Так или иначе, отдавая должное и тому, и другому, и третьему, можно заключить, что согласно излагаемой гипотезе, межядерные взаимодействия в живой клетке вполне возможны. И не будет в том помехой кулоновский барьер: природа сумела обойти этот барьер без высоких энергий и температур, мягко и нежно, КОРРЕСПОНДЕНТ. Вы считаете, что в живой клетке возникает вихревое электромагнитное поле. Оно удерживает протоны как бы в своей сетке и разгоняет их, ускоряет. Поле это излучают, генерируют электроны атомов железа. Есть группы из четырех таких атомов. Они называются у специалистов так: гемы. Железо в них двух- и трехвалентно. И обе эти формы обмениваются электронами, перескоки которых и порождают поле. Частота его невероятно велика, по вашей оценке 1028 герц. Она намного превосходит частоту видимого света, порождаемого обычно тоже перескоками электронов с одного атомного уровня на другой. Не считаете ли вы, что эта оценка частоты поля в клетке вами очень завышена? Г.Н. ПЕТРАКОВИЧ. Отнюдь нет. КОРРЕСПОНДЕНТ. Ваш ответ мне понятен. Ведь именно очень высокие частоты и соответствующие им малые длины волн связаны с большой энергией квантов. Так, ультрафиолет с его короткими волнами действует сильнее, чем обычные лучи света. Для разгона протонов нужны очень _ короткие волны. Возможны ли проверки самой схемы ускорения протонов и частоты внутриклеточного поля? Г.Н. ПЕТРАКОВИЧ. Итак, открытие: в митохондриях клеток генерируется сверхвысокочастотный, сверхкоротковолновый переменный электрический ток и по законам физики, соответственно ему - сверхкоротковолновое и сверхвысокочастотное переменное электромагнитное поле. Самое коротковолновое и самое высокочастотное из всех переменных электромагнитных полей в природе. Еще не созданы приборы, которыми можно было бы измерить такую высокую частоту и такую короткую волну, поэтому таких полей пока для нас как бы вовсе не существует. И открытия пока что не существует... Тем не менее вновь обратимся к законам физики. По этим законам точечные переменные электромагнитные поля самостоятельно не существуют, они мгновенно, со скоростью света сливаются между собой путем синхронизации и резонанса, значительно увеличивающим напряжение такого поля. Сливаются точечные электромагнитные поля, образуемые в электромагнитике перемещающимися электронами, далее сливаются все поля уже митохондрии. Образуется объединенное сверхвысокочастотное, сверхкоротковолновое переменное поле для всей митохондрии. В этом поле и удерживаются протоны. Но митохондрии в одной клетке не две и не три - в каждой клетке их насчитывается десятки, сотни, а в некоторых - даже тысячи, и в каждой из них образуется это сверхкоротковолновое поле; и эти поля устремляются к слиянию между собой, все с той же синхронизацией и эффектом резонанса, но уже во всем пространстве клетки - в цитоплазме. Вот это стремление переменного электромагнитного поля митохондрии к слиянию с другими такими же полями в цитоплазме есть та самая "тягловая сила", та энергия, что с ускорением "выбрасывает" протоны из митохондрии в пространство клетки. Так срабатывает внут-римитохондриальный "синхрофазотрон". Следует помнить, что протоны движутся к ядрам атомов-мишеней в клетке в значительно усиленном поле - настолько коротковолновом, что оно легко, как по волноводу, пройдет между ближайшими атомами даже в металлической решетке. Это поле легко "пронесет" с собой протон, размеры которого в сотню тысяч раз меньше любого атома, и настолько высокочастотно, что оно нисколько не потеряет при этом своей энергии. Такое обладающее сверхпроницаемостью поле возбудит и те протоны, которые входят в состав ядра атома-мишени. И главное - это поле приблизит к ним "налетающий" протон настолько, что позволит этому "налетающему" отдать ядру часть своей кинетической энергии. Самое большое количество энергии выделяется при альфа-распаде. При этом из ядра с огромной скоростью выбрасываются альфа-частицы, представляющие собой прочно связанные два протона и два нейтрона (то есть ядра атомов гелия). В отличие от ядерного взрыва при "холодном термояде" в зоне реакции не происходит накопления критической массы. Распад или синтез могут немедленно прекратиться. Не наблюдается радиации, поскольку альфа-частицы вне электромагнитного поля немедленно превращаются в атомы гелия, а протоны - в молекулярный водород, воду или перекиси. В то же время организм способен сам себе путем "холодного термояда" создавать необходимые ему химические элементы из других химических элементов, нейтрализовать вредные для него вещества. В зоне свершения "холодного термояда" формируются голо граммы, отражающие взаимодействия протонов с ядрами атомов-мишеней. В конечном итоге эти голограммы в неискаженном виде выносятся электромагнитными полями в ноосферу и становятся основой энергоинформационного поля ноосферы. Человек способен произвольно, с помощью электромагнитных линз, роль которых в живом организме выполняют молекулы-пьезокристаллы, фокусировать энергию протонов, и особенно альфа-частиц, в мощные пучки. При этом демонстрируя потрясающие воображение феномены: поднятие и передвижение неимоверных тяжестей, хождение по раскаленным камням и углям, левитацию, телепортацию, телекинез и многое другое. Не может такого быть, чтобы в мире все исчезало бесследно, наоборот, следует думать, что существует некий всемирный "банк", всемирное биополе, с которым сливались и сливаются поля всех живших и живущих на Земле. Это биополе может быть представлено сверхмощным, сверхвысокочастотным, сверхкоротковолновым и сверхпроникающим переменным электромагнитным полем вокруг Земли (и тем самым - вокруг и через нас). В этом поле в идеальном порядке удерживаются ядерные заряды протонных голографических "фильмов" о каждом из нас - о людях, о бактериях и слонах, о червяках, о траве, планктоне, саксауле, живших когда-то и живущих ныне. Живущие ныне и поддерживают энергией своего поля это биополе. Но только редкие единицы имеют доступ к его информационным сокровищам. Это память планеты, ее биосферы. Непознанное еще всемирное биополе обладает колоссальной, если не беспредельной, энергией, все мы купаемся в океане этой энергии, но не чувствуем ее, как не чувствуем окружающий нас воздух, и потому не чувствуем, что она вокруг нас есть... Роль ее будет возрастать. Это наш резерв, наша поддержка. КОРРЕСПОНДЕНТ. Само по себе это поле планеты, однако, не заменит рабочие руки и творческий ум. Оно лишь создает предпосылки для проявления человеческих способностей.
 Г.Н. ПЕТРАКОВИЧ.
Еще один аспект темы. Наши глаза, если и не зеркало души, то прозрачные их среды -зрачок и радужка - все же являются экранами для постоянно исходящего из нас топографического "кино". Через зрачки пролетают "цельные" голограммы, а в радужках протоны, несущие в себе значительный заряд кинетической энергии, непрерывно возбуждают молекулы в глыбках пигмента. Они будут возбуждать их до тех пор, пока в клетках, "пославших" к этим молекулам свои протоны, будет все в порядке. Погибнут клетки, еще что-то случится с ними, с органом - тотчас изменится структура в глыбках пигментов. Это четко зафиксируют опытные иридодиагносты: они уже точно - по проекциям в радужке - знают, какой орган заболел и даже чем. Ранняя и точная диагностика! Некоторые медики не очень благосклонно относятся к своим коллегам-иридодиагностам, считая их чуть ли не шарлатанами. Напрасно! Иридодиагностике, как простому, общедоступному, дешевому, легко переводимому на математический язык, а главное - точному и раннему методу диагностики различных болезней уже в ближайшем будущем светит "зеленый свет". Единственным недостатком метода было отсутствие теоретической базы. Фундамент ее изложен выше. КОРРЕСПОНДЕНТ.
Думаю, для наших читателей нужно бы пояснить процесс образования голограмм каждого индивида. Вы это сделаете лучше меня. Г.Н. ПЕТРАКОВИЧ.
Представим себе взаимодействия ускоренных протонов с какой-либо крупной объемной (трехмерной) молекулой в клетке, происходящие очень быстро. На такие взаимодействия с ядрами атомов-мишеней, составляющих эту крупную молекулу, будет израсходовано множество протонов, что оставит, в свою очередь, в пучке протонов тоже объемный, но "негативный" след в виде вакуума, "дырок". Этот след и будет самой настоящей голограммой, воплотившей в себе и сохранившей часть прореагировавшей с протонами структуры самой молекулы. Серия голограмм (что и происходит "в натуре") отобразит и сохранит не только физический "облик" молекулы, но и порядок физических и химических превращений отдельных ее частей и всей молекулы в целом за определенный промежуток времени. Такие голограммы, сливаясь в более крупные объемные изображения, могут отобразить жизненный цикл всей клетки, множества соседних клеток, органов и частей тела - всего тела. Есть еще одно следствие. Вот оно. В живой природе, независимо от сознания, мы общаемся прежде всего полями. При таком общении, войдя в резонанс с другими полями, мы рискуем утратить, частично или полностью, свою индивидуальную частоту (как и чистоту), и если в общении с зеленой природой это означает "раствориться в природе", то в общении с людьми, особенно с теми, кто обладает сильным полем, это значит частично или полностью утратить свою индивидуальность - стать "зомби" (по Тодору Дичеву). Технических аппаратов "зомбирования" по программе нет и вряд ли они когда-либо будут созданы, но воздействия одного человека на другого в этом плане вполне возможны, хотя, с позиций морали, недопустимы. Оберегая себя, над этим следует задуматься, особенно когда дело касается шумных коллективных действий, в которых всегда преобладает не разум и даже не истинное чувство, но фанатизм - печальное дитя злонамеренного резонанса. Поток протонов может только увеличиваться за счет слияния с другими потоками, но никак, в противовес, например, электронному потоку, не смешиваться - и тогда он может нести в себе полную информацию уже о целых органах и тканях, в том числе - и о таком специфическом органе, как мозг. По-видимому, мы мыслим программами, и эти голограммы способны передавать потоком протонов через взгляд - тому доказательство не только "выразительность" нашего взгляда, но и то, что животные способны усваивать наши голограммы. В подтверждение этому можно сослаться на опыты известного дрессировщика В.Л. Дурова, в которых принимал участие и академик В.М. Бехтерев. В этих опытах собакам специальной комиссией сиюминутно придумывались какие-либо посильные им задания, В.Л. Дуров тут же "гипнотическим взглядом" передавал собакам эти задания (при этом, как он говорил, он сам как бы становился "собакой" и вместе с ними мысленно выполнял задания), и собаки в точности выполняли все предписания комиссии. Кстати, и фотографирование галлюцинаций можно связать с голографическим мышлением и передачей образов потоком протонов через взгляд. Очень важный момент: несущие информацию протоны своей энергией "метят" белковые молекулы своего тела, при этом каждая "меченая" молекула приобретает свой собственный спектр, и этим спектром она отличается от точно такой же по химическому составу молекулы, но принадлежащей "чужому" телу. Принцип несовпадения (или совпадения) по спектру молекул белка лежит в основе иммунных реакций организма, воспаления, а также тканевой несовместимости, о чем мы уже упоминали. Механизм обоняния тоже построен на принципе спектрального анализа возбужденных протонами молекул. Но в этом случае протонами облучаются все находящиеся во вдыхаемом через нос воздухе молекулы вещества с мгновенным анализом их спектра (механизм очень близок к механизму цветоощущения). Но есть "работа", которую выполняет только высокочастотное переменное электромагнитное поле - это работа "второго", или "периферического", сердца, о котором в свое время много писали, но механизм которого еще никто не раскрыл. Это особая тема для разговора. Продолжение следует...
Г.Н. ПЕТРАКОВИЧ.
Еще один аспект темы. Наши глаза, если и не зеркало души, то прозрачные их среды -зрачок и радужка - все же являются экранами для постоянно исходящего из нас топографического "кино". Через зрачки пролетают "цельные" голограммы, а в радужках протоны, несущие в себе значительный заряд кинетической энергии, непрерывно возбуждают молекулы в глыбках пигмента. Они будут возбуждать их до тех пор, пока в клетках, "пославших" к этим молекулам свои протоны, будет все в порядке. Погибнут клетки, еще что-то случится с ними, с органом - тотчас изменится структура в глыбках пигментов. Это четко зафиксируют опытные иридодиагносты: они уже точно - по проекциям в радужке - знают, какой орган заболел и даже чем. Ранняя и точная диагностика! Некоторые медики не очень благосклонно относятся к своим коллегам-иридодиагностам, считая их чуть ли не шарлатанами. Напрасно! Иридодиагностике, как простому, общедоступному, дешевому, легко переводимому на математический язык, а главное - точному и раннему методу диагностики различных болезней уже в ближайшем будущем светит "зеленый свет". Единственным недостатком метода было отсутствие теоретической базы. Фундамент ее изложен выше. КОРРЕСПОНДЕНТ.
Думаю, для наших читателей нужно бы пояснить процесс образования голограмм каждого индивида. Вы это сделаете лучше меня. Г.Н. ПЕТРАКОВИЧ.
Представим себе взаимодействия ускоренных протонов с какой-либо крупной объемной (трехмерной) молекулой в клетке, происходящие очень быстро. На такие взаимодействия с ядрами атомов-мишеней, составляющих эту крупную молекулу, будет израсходовано множество протонов, что оставит, в свою очередь, в пучке протонов тоже объемный, но "негативный" след в виде вакуума, "дырок". Этот след и будет самой настоящей голограммой, воплотившей в себе и сохранившей часть прореагировавшей с протонами структуры самой молекулы. Серия голограмм (что и происходит "в натуре") отобразит и сохранит не только физический "облик" молекулы, но и порядок физических и химических превращений отдельных ее частей и всей молекулы в целом за определенный промежуток времени. Такие голограммы, сливаясь в более крупные объемные изображения, могут отобразить жизненный цикл всей клетки, множества соседних клеток, органов и частей тела - всего тела. Есть еще одно следствие. Вот оно. В живой природе, независимо от сознания, мы общаемся прежде всего полями. При таком общении, войдя в резонанс с другими полями, мы рискуем утратить, частично или полностью, свою индивидуальную частоту (как и чистоту), и если в общении с зеленой природой это означает "раствориться в природе", то в общении с людьми, особенно с теми, кто обладает сильным полем, это значит частично или полностью утратить свою индивидуальность - стать "зомби" (по Тодору Дичеву). Технических аппаратов "зомбирования" по программе нет и вряд ли они когда-либо будут созданы, но воздействия одного человека на другого в этом плане вполне возможны, хотя, с позиций морали, недопустимы. Оберегая себя, над этим следует задуматься, особенно когда дело касается шумных коллективных действий, в которых всегда преобладает не разум и даже не истинное чувство, но фанатизм - печальное дитя злонамеренного резонанса. Поток протонов может только увеличиваться за счет слияния с другими потоками, но никак, в противовес, например, электронному потоку, не смешиваться - и тогда он может нести в себе полную информацию уже о целых органах и тканях, в том числе - и о таком специфическом органе, как мозг. По-видимому, мы мыслим программами, и эти голограммы способны передавать потоком протонов через взгляд - тому доказательство не только "выразительность" нашего взгляда, но и то, что животные способны усваивать наши голограммы. В подтверждение этому можно сослаться на опыты известного дрессировщика В.Л. Дурова, в которых принимал участие и академик В.М. Бехтерев. В этих опытах собакам специальной комиссией сиюминутно придумывались какие-либо посильные им задания, В.Л. Дуров тут же "гипнотическим взглядом" передавал собакам эти задания (при этом, как он говорил, он сам как бы становился "собакой" и вместе с ними мысленно выполнял задания), и собаки в точности выполняли все предписания комиссии. Кстати, и фотографирование галлюцинаций можно связать с голографическим мышлением и передачей образов потоком протонов через взгляд. Очень важный момент: несущие информацию протоны своей энергией "метят" белковые молекулы своего тела, при этом каждая "меченая" молекула приобретает свой собственный спектр, и этим спектром она отличается от точно такой же по химическому составу молекулы, но принадлежащей "чужому" телу. Принцип несовпадения (или совпадения) по спектру молекул белка лежит в основе иммунных реакций организма, воспаления, а также тканевой несовместимости, о чем мы уже упоминали. Механизм обоняния тоже построен на принципе спектрального анализа возбужденных протонами молекул. Но в этом случае протонами облучаются все находящиеся во вдыхаемом через нос воздухе молекулы вещества с мгновенным анализом их спектра (механизм очень близок к механизму цветоощущения). Но есть "работа", которую выполняет только высокочастотное переменное электромагнитное поле - это работа "второго", или "периферического", сердца, о котором в свое время много писали, но механизм которого еще никто не раскрыл. Это особая тема для разговора. Продолжение следует...
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Основным источником энергии для всех живых существ, населяющих нашу планету, служит энергия солнечного света, которую используют непосредственно только клетки зеленых растений, водорослей, зеленых и пурпурных бактерий. В этих клетках из углекислого газа и воды в процессе фотосинтеза образуются органические вещества (углеводы, жиры, белки, нуклеиновые кислоты и др.). Поедая растения, животные получают органические вещества в готовом виде. Энергия, запасенная в этих веществах, переходит вместе с ними в клетки гетеротрофных организмов.
В клетках животных организмов энергия органических соединений при их окислении превращается в энергию АТФ. (Углекислый газ и вода, выделяющиеся при этом, вновь используются автотрофными организмами для процессов фотосинтеза.) За счет энергии АТФ осуществляются все процессы жизнедеятельности: биосинтез органических соединений, движение, рост, деление клеток и др.
Аденозинтрифосфат - нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. АТФ был открыт в 1929 году Карлом Ломанном, а в1941 году Фриц Липман показал, что АТФ является основным переносчиком энергии в клетке .
По строению АТФ сходна с адениновым нуклеотидом, входящим в состав РНК, только вместо одной фосфорной кислоты в состав АТФ входят три остатка фосфорной кислоты. Клетки не в состоянии содержать кислоты в заметных количествах, а только их соли. Поэтому фосфорная кислота входит в АТФ в виде остатка (вместо ОН-группы кислоты имеется отрицательно заряженный атом кислорода).
Эти реакции обратимы, то есть АМФ может переходить в АДФ и далее в АТФ, аккумулируя энергию. Разрушение обычной пептидной связи высвобождает лишь 12 кДж/моль энергии. А связи, которыми присоединены остатки фосфорной кислоты, высокоэнергетичны (их еще называют макроэргическими): при разрушении каждой из них выделяется 40 кДж/моль энергии. Поэтому АТФ играет в клетках центральную роль универсального биологического аккумулятора энергии. Молекулы АТФ синтезируются в митохондриях и хлоропластах (лишь незначительное их количество синтезируется в цитоплазме), а затем поступают к различным органоидам клетки, обеспечивая энергией все процессы жизнедеятельности.
При усиленной нагрузке (например, в беге на короткие дистанции) мышцы работают исключительно за счет запаса АТФ. В клетках мышц этого запаса хватает на несколько десятков сокращений, а дальше количество АТФ должно восполняться. Синтез АТФ из АДФ и АМФ происходит за счет энергии, выделяющейся при расщеплении углеводов, липидов и других веществ. На выполнение умственной работы также затрачивается большое количество АТФ. По этой причине людям умственного труда требуется повышенное количество глюкозы, расщепление которой обеспечивает синтез АТФ .
АТФ является неустойчивой молекулой. В небуферизованной воде, он гидролизуется до АДФ и фосфата. Это потому, что прочность связей между фосфатными группами в АТФ меньше, чем прочность водородных связей (гидратации облигации), между своей продукции (АДФ + фосфат), и воду. Таким образом, если АТФ и АДФ находятся в химическом равновесии в воде, почти все АТФ будет в конечном счете превращается в AДФ. Система, которая далека от равновесия содержит свободную энергию Гиббса, и способен делать работу. Живые клетки поддерживают соотношение АТФ в АДФ в точке десять порядков от равновесия, при концентрации АТФ в тысячу раз выше, чем концентрация AДФ. Это смещение от положения равновесия означает, что гидролиз АТФ в клетке выпускает большое количество свободной энергии .
Две высокоэнергетические фосфатные связи (те, которые соединяют соседние фосфаты) в молекулы АТФ несут ответственность за высокое содержание энергии этой молекулы. Энергия, запасенная в АТФ может быть освобождена от гидролиза. Расположенный дальний от сахара рибозы, г-фосфатная группа имеет более высокую энергию гидролиза, чем любой б - или в-фосфат. Связи, образованные после гидролиза или фосфорилирования остатка АТФ - ниже по энергии, чем других связей АТФ. Во время фермент-катализируемого гидролиза АТФ или АТФ фосфорилирования, имеющиеся свободная энергия может быть использована на живые системы, чтобы сделать работу .
Любая нестабильная система потенциально реактивных молекул потенциально может служить способом хранения свободной энергии, если клетки сохранили свою концентрацию далеко от точки равновесия реакции. Однако, как и в случае с большинством полимерных биомолекул, разбивка РНК, ДНК и АТФ в простые мономеры обусловлена как выделение энергии и энтропии, увеличение соображения, как в стандартной концентрации, а также тех концентрациях, в которых встречается в клетке .
Стандартное количество энергии, выделяющейся в результате гидролиза АТФ может быть рассчитана по изменениям в энергии, не связанных с природными (стандартные) условия, то исправление биологической концентрации. Чистое изменение в тепловой энергии (энтальпии) при стандартной температуре и давлении разложения АТФ в АДФ и неорганических фосфатов - 20,5 кДж / моль, с изменением свободной энергии на 3,4 кДж / моль. Энергия выпускается путем расщепления фосфата или пирофосфата от АТФ на государственный стандарт 1 М являются:
Эти значения могут быть использованы для расчета изменения энергии в физиологических условиях и клеточных АТФ / АДФ. Тем не менее, более представительная значимость, называемая энергетическим зарядом, чаще работает. Значения приведены для свободной энергии Гиббса. Эти реакции зависят от ряда факторов, включая общую ионную силу и присутствие щелочно-земельных металлов, такие как ионы Mg 2 + и Ca 2 + . В обычных условиях, ДG составляет около - 57 кДж / моль (-14 ккал / моль) .
белок биологический аккумулятор энергия
Фосфорилирование АДФ возможно двумя способами: субстратное фосфорилирование и окислительное фосфорилирование (используя энергию окисляющихся веществ). Основная масса АТФ образуется на мембранах митохондрий в ходе окислительного фосфорилирования H-зависимой АТФ-синтазой. Субстратное фосфорилирование АТФ не требует участия мембранных ферментов, оно происходит в процессе гликолиза или путём переноса фосфатной группы с других макроэргических соединений.
В организме АТФ является одним из самых часто обновляемых веществ. Так у человека продолжительность жизни одной молекулы АТФ менее 1 мин. В течение суток одна молекула АТФ проходит в среднем 2000-3000 циклов ресинтеза (человеческий организм синтезирует около 40 кг АТФ в день), то есть запаса АТФ в организме практически не создаётся, и для нормальной жизнедеятельности необходимо постоянно синтезировать новые молекулы АТФ .
2 .1.1 Гл ико лиз - ферментативный процесс последовательного расщепления глюкозы в клетках, сопровождающийся синтезом АТФ. Гликолиз при аэробных условиях ведёт к образованию пировиноградной кислоты (пирувата), гликолиз в анаэробных условиях ведёт к образованию молочной кислоты (лактата). Гликолиз является основным путём катаболизма глюкозы в организме животных .
Процесс гликолиза условно можно разделить на два этапа. Первый этап, протекающий с расходом энергии 2-х молекул АТФ, заключается в расщеплении молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся синтезом АТФ. Сам по себе гликолиз является полностью анаэробным процессом, то есть не требует для протекания реакций присутствия кислорода.
У аэробных организмов конечные продукты гликолиза подвергаются дальнейшим превращениям в биохимических циклах, относящихся к клеточному дыханию. В итоге после полного окисления всех метаболитов одной молекулы глюкозы на последнем этапе клеточного дыхания - окислительном фосфорилировании, происходящем на митохондриальной дыхательной цепи в присутствии кислорода, - дополнительно синтезируются ещё 34 или 36 молекулы АТФ на каждую молекулу глюкозы.
Фосфорилирование глюкозы преследует две цели: во-первых, из-за того что плазматическая мембрана, проницаемая для нейтральной молекулы глюкозы, не пропускает отрицательно заряженные молекулы Г-6-Ф, фосфорилированная глюкоза оказывается запертой внутри клетки. Во-вторых, при фосфорилировании глюкоза переводится в активную форму, способную участвовать в биохимических реакциях и включаться в метаболические циклы.
Это первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1,3-дифосфоглицерат) вместо двух потраченных в реакциях 1 и 3 . Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
Гликолиз - катаболический путь исключительной важности. Он обеспечивает энергией клеточные реакции, в том числе и синтез белка. Промежуточные продукты гликолиза используются при синтезе жиров. Пируват также может быть использован для синтезааланина, аспартата и других соединений. Благодаря гликолизу производительность митохондрий и доступность кислорода не ограничивают мощность мышц при кратковременных предельных нагрузках .
На I стадии этого процесса пируват теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E 1). На II стадии оксиэтильная группа комплекса E 1 -ТПФ-СНОН-СН 3 окисляется с образованием ацетильнойгруппы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментомдигидроли-поилацетилтрансферазой (Е 2). Этот фермент катализирует III стадию - перенос ацетильнойгруппы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид-Е 2 . При участии фермента дигидролипоилдегидрогеназы (Е 3) осуществляется переносатомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН 2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н + .
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E 1 , ли-поамид-Е 2 и ФАД-Е 3), а два - легко диссоциируют (HS-KoA и НАД).
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано, что составляющие комплекс полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
2 .1.3 Цикл трикарбоновых кисло т (цикл Кре бса , цитра тный цикл ) - центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух - и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO 2 . При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии - АТФ.
Цикл Кребса - это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др.
У эукариот все реакции цикла Кребса протекают внутри митохондрий, причём катализирующие их ферменты, кроме одного, находятся в свободном состоянии в митохондриальном матриксе, исключение составляет сукцинатдегидрогеназа, которая локализуется на внутренней митохондриальной мембране, встраиваясь в липидный бислой. У прокариот реакции цикла протекают в цитоплазме .
Цикл Кребса регулируется "по механизму отрицательной обратной связи", при наличии большого количества субстратов (ацетил-КоА, оксалоацетат), цикл активно работает, а при избытке продуктов реакции (НАД, ATФ) тормозится. Регуляция осуществляется и при помощи гормонов, основным источником ацетил-КоА является глюкоза, поэтому гормоны, способствующие аэробному распаду глюкозы, способствуют работе цикла Кребса. Такими гормонами являются:
· б-субъединица транспортирует АТФ к мембранному переносчику, который "выбрасывает" АТФ в цитоплазму. Взамен из цитоплазмы этот же переносчик транспортирует АДФ. На внутренней мембране митохондрий также находится переносчик Фосфата из цитоплазмы в митохондрию, но для его работы необходим протон водорода. Такие переносчики называются транслоказами .
Субстр атное фосфорил ирование (биохимическое), синтез богатых энергией фосфорных соединений за счёт энергии окислительно-восстановительных реакций гликолиза (катализируемых фосфоглице-ральдегиддегидрогеназой и енолазой) и при окислении a-кетоглутаровой кислоты в цикле трикарбоновых кислот (под действием a-кетоглутаратдегидрогеназы и сукцинаттиокиназы). Для бактерий описаны случаи С. ф. при окислении пировиноградной кислоты.С. ф., в отличие от фосфорилирования в цепи переноса электронов, не ингибируется "разобщающими" ядами (например, динитрофенолом) и не связано с фиксацией ферментов в мембранах митохондрий. Вклад С. ф. в клеточный фонд АТФ в аэробных условиях значительно меньше, чем вклад фосфорилирования в цепи переноса электронов .
Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. Всё это реакции синтеза сложных веществ в организме: осуществление активного переноса молекул через биологические мембраны, в том числе и для создания трансмембранного электрического потенциала; осуществления мышечного сокращения .
АТФ представляет собой термодинамически нестойкое соединение. Нестабильность АТФ определяется, во - первых, электростатическим отталкиванием в области кластера одноимённых отрицательных зарядов, что приводит к напряжению всей молекулы, однако сильнее всего связи - Р - О - Р, и во - вторых, конкретным резонансом. В соответствии с последним фактором существует конкуренция между атомами фосфора за неподелённые подвижные электроны атома кислорода, расположенного между ними, поскольку на каждом атоме фосфора имеется частичный положительный заряд в следствии значительного электронаицепторного влияния групп Р=О и Р - О-. Таким образом, возможность существования АТФ определяется наличием достаточного количества химической энергии в молекуле, позволяющей компенсировать эти физико - химические напряжения. В молекуле АТФ имеется две фосфоангидридных (пирофосфатных) связи, гидролиз которых сопровождается значительным уменьшением свободной энергии (при рН 7,0 и 37 о С).
Фосфорилирование АДФ является эндергоническим процессом и требует источника энергии. Как отмечалось ранее, в природе преобладает два таких источника энергии - это солнечная энергия и химическая энергия восстановленных органических соединений. Зелёные растения и некоторые микроорганизмы способны трансформировать энергию, поглощённых квантов света в химическую энергию, которая расходуется на фосфорилирование АДФ в световой стадии фотосинтеза. Этот процесс регенерации АТФ получил название фотосинтетического фосфорилирования. Трансформация энергии окисления органических соединений в макроэнергетические связи АТФ в аэробных условиях происходит преимущественно путём окислительного фосфорилирования. Свободная энергия, необходимая для образования АТФ, генерируется в дыхательной окислительной цепи митаходрий.
Известен ещё один тип синтеза АТФ, получивший название субстратного фосфорилирования. В отличии от окислительного фосфорилирования, сопряжённого с переносом электронов, донором активированной фосфорильной группой (- РО3 Н2), необходимой для регенерации АТФ, являются интермедианты процессов гликолиза и цикла трикарбоновых кислот. Во всех этих случаях окислительные процессы приводят к образованию высокоэнергетических соединений: 1,3 - дифосфоглицерата (гликолиз), сукцинил - КоА (цикл трикарбоновых кислот), которые при участии соответствующих ферментов способны фолирировать АДФ и образовывать АТФ. Трансформация энергии на уровне субстрата является единственным путём синтеза АТФ в анаэробных организмах. Этот процесс синтеза АТФ позволяет поддерживать интенсивную работу скелетных мышц в периоды кислородного голодания. Следует помнить, что он является единственным путём синтеза АТФ в зрелых эритроцитах не имеющих митохондрий.
где Ф - фермент, Е - освобождающая энергия. В этой реакции образуется аденозинфосфорная кислота (АДФ) - остаток молекулы АТФ и органический фосфат. Энергию АТФ все клетки используют для процессов биосинтеза, движения, производство тепла, нервных импульсов, свечений (например, улюминисцентных бактерий), то есть для всех процессов жизнедеятельности .
Запас АТФ в клетке невелик. Так, в мышце запаса АТФ хватает на 20 - 30 сокращений. При усиленной, но кратковременной работе мышцы работают исключительно за счёт расщепления содержащейся в них АТФ. После окончания работы человек усиленно дышит - в этот период происходит расщепление углеводов и других веществ (происходит накопление энергии) и запас АТФ в клетках восстанавливается.
Живая клетка является далека от равновесия химическая система: ведь приближение живой системы к равновесию означает ее распад и смерть. Продукт каждого фермента обычно быстро расходуется, поскольку используется в качестве субстрата другим ферментом данного метаболического пути. Еще более важно, что большое количество ферментативных реакций связана с расщеплением АТФ на АДФ и неорганический фосфат. Чтобы это было возможным, пул АТФ в свою очередь должен поддерживаться на уровне, далеком от равновесия, так чтобы отношение концентрации АТФ в концентрации продуктов его гидролиза было высоким. Таким образом, пул АТФ играет роль "аккумулятора", поддерживающий постоянный перенес в клетке энергии и атомов по метаболическим путям, определяются присутствовать ферментами.
На пути от А - Н и Б - ОН-А - Б, связанном с гидролизом АТФ, энергия гидролиза сначала переводит Б - ОН в высокоэнергетическую промежуточную соединение, которое затем непосредственно реагирует с А - Н, образуя А - Б. простой механизм данного процесса включает в себя перенес фосфата от АТФ к Б - ОН с образованием Б - ОРО 3 , или Б - О - Р, причем в этом случае суммарная реакция происходит лишь в две стадии:
Величина G гидролиза АТФ до АДФ и неорганического фосфата зависит от концентрации всех реагирующих веществ и обычно для клетки условиях лежит в пределах от - 11 до - 13 ккал / моль. Реакция гидролиза АТФ, наконец, может быть использована для осуществления термодинамически невыгодной реакции со значением G, равным примерно +10 ккал / моль, конечно, в присутствии соответствующей последовательности реакций. Однако для многих реакций биосинтеза оказывается недостаточным даже? G = - 13 ккал / моль. В этих и других случаях путь гидролиза АТФ изменяется таким образом, что сначала образуются АМФ и РР (пирофосфат). На следующей стадии пирофосфат также подвергается гидролизу; общее изменение свободной энергии всего процесса составляет примерно - 26 ккал / моль.
Каким образом энергия гидролиза пирофосфата используется в биосинтетических реакциях? Один из путей можно продемонстрировать на примере приведенного выше синтеза соединения А - Б с А - Н и Б - ОН. При помощи соответствующего фермента Б - ОН может вступить в реакцию с АТФ и превратиться в высокоэнергетическую соединение Б - О - Р - Р. Теперь реакция состоит из трех стадий:
Поскольку фермент всегда ускоряет катализовану ним реакцию как в прямом, так и в обратном направлении, соединение А - Б может распадаться, реагируя с пирофосфатом (реакция, обратная стадии 2). Однако энергетически выгодная реакция гидролиза пирофосфата (стадия 3) способствует поддержанию стабильности соединения А-Б за счет того, что концентрация пирофосфата остается очень низкой (это предотвращает протекание реакции, обратной к стадии 2). Таким образом, энергия гидролиза пирофосфата обеспечивает протекание реакции в прямом направлении. Примером важной биосинтетических реакции такого типа является синтез полинуклеотидов .
Во всех известных организмах, дезоксирибонуклеотиды, которые составляют ДНК синтезируется под действием ферментов рибонуклеотид-редуктазы (РНР) на соответствующих рибонуклеотидах. Эти ферменты снижают остатка сахара отрибозы, чтобы дезоксирибоза путем удаления кислорода из 2" гидроксильные группы, субстраты рибонуклеозид дифосфаты и продукты дезоксирибонуклеозид дифосфаты. Все ферменты редуктазы использоют общие сульфгидрильные радикального механизма зависит от реактивного цистеина остатков, которые окисляются с образованием дисульфидных связей в ход реакции. РНР фермент перерабатывается в результате реакции с тиоредоксином или глутаредоксином.
При синтезе нуклеиновых кислот РНК, аденозина, полученных от АТФ является одним из четырех нуклеотидов включены непосредственно в молекулах РНК, РНК-полимеразы. Энергия, это полимеризация происходит с отщеплением пирофосфата (две фосфатные группы). Этот процесс аналогичен в биосинтезе ДНК, за исключением того, что АТФ сводится к дезоксирибонуклеотиду дАТФ, перед включением в ДНК .
АТФ используется для многих клеточных функций, включая транспортную работу перемещения веществ через клеточные мембраны. Он также используется для механической работы, поставка энергии, необходимой для мышечного сокращения. Она поставляет энергию не только в сердечной мышце (для циркуляции крови) и скелетных мышц (например, за грубое движение тела), но также и к хромосомам и жгутиков, чтобы они могли выполнять свои многочисленные функции. Большую роль АТФ в химической работе, предоставлению необходимой энергии для синтеза нескольких тысяч типов макромолекул, что клетка должна существовать.
АТФ также используется в качестве включения-выключения как для контроля химических реакций и для отправки информации. Форма белковых цепей, которые производят строительные блоки и другие структуры, используемые в жизни определяется в основном слабые химические связи, которые легко исчезают и переструктурируются. Эти цепи могут сократить, удлинить, а также изменить форму в ответ на ввод или вывод энергии. Изменения в цепях изменить форму белка, а также может изменять свои функции или вызвать его, чтобы стать активным или неактивным.
Таким образом, АТФ передает энергию между пространственно разделенных метаболических реакций. АТФ является основным источником энергии для большинства клеточных функций. Это включает в себя синтез макромолекул, включая ДНК и РНК, и белки. АТФ также играет важную роль в транспорте макромолекул через клеточные мембраны, например, экзоцитоза и эндоцитоза.
ATФ участвует в поддержании клеточной структуры путем облегчения монтажа и демонтажа элементов цитоскелета. В связи с этим процессом, АТФ, необходимых для сокращения нитей актина и миозина необходимых для мышечного сокращения. Этот последний процесс является одним из основных требований энергию животных и имеет важное значение для движения и дыхания.
АТФ имеет решающее значение в передаче сигнала процессов. Он используется киназ в качестве источника фосфатных групп в их реакции фосфата передачи. Киназы на подложках, таких как белки или липиды мембраны являются распространенной формой сигнала. Фосфорилирование белка по киназе могут активировать этот каскад, такие как митогенактивированной протеинкиназыкаскада.
1. Аденозинтрифосфат - нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. Химически АТФ представляет собой трифосфорный эфир аденозина, который является производным аденина и рибозы. По строению АТФ сходна с адениновым нуклеотидом, входящим в состав РНК, только вместо одной фосфорной кислоты в состав АТФ входят три остатка фосфорной кислоты. Клетки не в состоянии содержать кислоты в заметных количествах, а только их соли. Поэтому фосфорная кислота входит в АТФ в виде остатка (вместо ОН-группы кислоты имеется отрицательно заряженный атом кислорода).
3. Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. В биоэнергетике живых организмов имеют значение: химическая энергия запасается путём образования АТФ, сопряжённого с экзергоническими катаболическими реакциями окисления органических субстратов; химическая энергия утилизируется путём расщепления АТФ, сопряжённого с эндергоническими реакциями анаболизма и другими процессами, требующими затраты энергии.
4. При усиленной нагрузке (например, в беге на короткие дистанции) мышцы работают исключительно за счет запаса АТФ. В клетках мышц этого запаса хватает на несколько десятков сокращений, а дальше количество АТФ должно восполняться. Синтез АТФ из АДФ и АМФ происходит за счет энергии, выделяющейся при расщеплении углеводов, липидов и других веществ. На выполнение умственной работы также затрачивается большое количество АТФ. По этой причине людям умственного труда требуется повышенное количество глюкозы, расщепление которой обеспечивает синтез АТФ.
1. Лемеза, Н.А. Пособие по биологии для поступающих в ВУЗы / Л.В. Камлюк Н.Д. Лисов. - Мн.: Юнипресс, 2011 г. - 624 с.
2. Lodish, H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J. Molecular Cell Biology, 5th ed. - New York: WH Freeman, 2004.
3. Романовский, Ю.М. Молекулярные преобразователи энергии живой клетки. Протонная АТФ-синтаза - вращающийся молекулярный мотор / Ю.М. Романовский А.Н. Тихонов // УФН. - 2010. - Т.180. - С.931 - 956.
4. Voet D, Voet JG. Biochemistry Vol 1 3rd ed. - Wiley: Hoboken, NJ. - N-Y: W. H. Freeman and Company, 2002. - 487 р.
5. Общая химия. Биофизическая химия. Химия биогенных элементов. М.: Высшая школа, 1993 г
6. Вершубский, А.В. Биофизика. / А.В. Вершубский, В.И. Прик-лонский, А.Н. Тихонов. - М: 471-481.
7. Альбертс Б. Молекулярная биология клетки в 3-х томах. / Альбертс Б., Брей Д., Льюис Дж. и др. М.: Мир, 1994.1558 с.
8. Николаев А.Я. Биологическая химия - М.: ООО "Медицинское информационное агенство", 1998.
9. Berg, J. M. Biochemistry, international edition. / Berg, J. M, Tymoczko, J. L, Stryer, L. - New York: WH Freeman, 2011; p 287.
10. Кнорре Д.Г. Биологическая химия: Учеб. для хим., биол. И мед. спец. вузов. - 3-е изд., испр. / Кнорре Д.Г., Мысина С.Д. - М.: Высш. шк., 2000. - 479 с.: ил.
11. Элиот, В. Биохимия и молекулярная биология / В. Элиот, Д. Элиот. - М.: Изд-во НИИ Биомедицинской химии РАМН, ООО "Материк-альфа", 1999, - 372 с.
12. Shina CL, K., 7 Areieh, W. On the Energetics of ATP Hydrolysis in Solution. Journal Of Physical Chemistry B,113 (47), (2009).
13. Berg, J. M. Biochemistry / J. M. Berg: J. L. Tymoczko, L. Stryer. - N-Y: W. H. Freeman and Company, 2002. - 1514 p.
...Органические соединения в организме человека. Строение, функции и классификация белков. Нуклеиновые кислоты (полинуклеотиды), особенности строений и свойства РНК н ДНК. Углеводы в природе и организме человека. Липиды - жиры и жироподобные вещества.
реферат , добавлен 06.09.2009
Процесс синтеза белков и их роль в жизнедеятельности живых организмов. Функции и химические свойства аминокислот. Причины их нехватки в организме человека. Виды продуктов, в которых содержатся незаменимые кислоты. Аминокислоты, синтезируемые в печени.
презентация , добавлен 23.10.2014
Энергетическая, запасающая и опорно-строительная функции углеводов. Свойства моносахаридов как основного источника энергии в организме человека; глюкоза. Основные представители дисахаридов; сахароза. Полисахариды, образование крахмала, углеводный обмен.
доклад , добавлен 30.04.2010
Функции обмена веществ в организме: обеспечение органов и систем энергией, вырабатываемой при расщеплении пищевых веществ; превращение молекул пищевых продуктов в строительные блоки; образование нуклеиновых кислот, липидов, углеводов и других компонентов.
реферат , добавлен 20.01.2009
Роль и значение белков, жиров и углеводов для нормального протекания всех жизненно важных процессов. Состав, структура и ключевые свойства белков, жиров и углеводов, их важнейшие задачи и функции в организме. Основные источники данных пищевых веществ.
презентация , добавлен 11.04.2013
Характеристика структуры холестериновых молекул как важного компонента клеточной мембраны. Исследование механизмов регуляции обмена холестерина в организме человека. Анализ особенностей возникновения избытка липопротеидов низкой плотности в кровотоке.
реферат , добавлен 17.06.2012
Обмен белков, липидов и углеводов. Типы питания человека: всеядность, раздельное и низкоуглеводное питание, вегетарианство, сыроедение. Роль белков в обмене веществ. Недостаток жиров в организме. Изменения в организме в результате изменения типа питания.
курсовая работа , добавлен 02.02.2014
Рассмотрение участия железа в окислительных процессах и в синтезе коллагена. Ознакомление со значением гемоглобина в процессах кровообразования. Головокружения, одышка и нарушение обмена веществ как результат дефицита железа в человеческом организме.
презентация , добавлен 08.02.2012
Свойства фтора и железа. Суточная потребность организма. Функции фтора в организме, влияние, смертельная доза, взаимодействие с другими веществами. Железо в организме человека, его источники. Последствия дефицита железа для организма и его переизбытка.
презентация , добавлен 14.02.2017
Белки как источники питания, их основные функции. Аминокислоты, участвующие в создании белков. Строение полипептидной цепи. Превращения белков в организме. Полноценные и неполноценные белки. Структура белка, химические свойства, качественные реакции.
| Статьи по теме: | |
|
Функции и графики. Понятие функции. Способы задания функции Табличный способ задания функции
Что означают слова "задать функцию"? Они означают: объяснить всем... "Духота и трупный запах": детские воспоминания о Сталинградской битве Воспоминания участников сталинградской битвы читать
В начале 70-х годов поздней осенью, когда уже выпал первый снег, мне... Трубецкой дети. Биографии декабристок. Биография Екатерины
Ивановны Трубецкой
План Введение 1 Биография 2 Жена декабриста 3 Произведения,... | |